Prehistory and Protohistory in Sicily. A Geometric Morphometrics Approach to Study the Biological History of Early Human Peopling of the Island
Gabriele Lauria
 https://orcid.org/0000-0002-0122-7093
https://orcid.org/0000-0002-0122-7093
Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, Via Archirafi 18, 90123 Palermo, Italy
Luca Sineo
https://orcid.org/0000-0001-8634-2295
Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, Via Archirafi 18, 90123 Palermo, Italy
Abstract. In recent years, the debate about the early human peopling of Sicily has almost exclusively focused on the archaeological evidence. The dispersal patterns and the possibility for, and degree of, admixture caused by ancient migratory flows have been only investigated in limited anthropological studies conducted on a short time spans. Recent craniofacial morphometric analyses that considered migratory flows and population influx have provided a more comprehensive approach. These analyses go beyond archaeologically based settlement hypotheses by merging previous archaeological evaluations and paleoclimatic studies with an anthropological approach. This study expands upon earlier morphometric work and provides an overview covering the period from the Upper Palaeolithic to the Iron Age. For this study, human skulls from Sicilian Prehistory and Protohistory were considered. These skulls were divided into six periods based on the dating of associated archaeological artifacts. Sample selection was based on a detailed bibliographic review of previously published archaeological and historical works. With the aim of associating the diachronic changes in cranial morphology with population migrations and admixture we performed a 3D geometric morphometrics (GM) comparative analysis. The data reported demonstrate that the first colonization of Sicily started during the Upper–Paleolithic when stable climatic conditions allowed hunter-gatherers to move from the Italian Peninsula to the island. Moreover, the results show a cyclical occupation of the land counterclockwise direction completed only with the hinterland colonization in the first historical periods with Greek and Punic colonization.
Keywords: geometric morphometrics, skulls, biological history, human peopling, Sicily
Introduction
The biological history of the human peopling of Sicily presents many interesting aspects. Its proximity to both the Italian Peninsula and North Africa and its unique physical geography have made the island a crossing point for several living species (e.g., Leithia melitensis, Aquila fasciata, Chamaerops humilis, Pinus halepensis etc.) (Bosch 2010; Massa 2011; Surdi 2011, Pignatti 2011). Sicily is the largest island in the Mediterranean Sea and is itself divided into a mosaic of habitats (due to coastlines, sea valleys, small cliffs, hills, and mountain ranges), each with its own microclimate (Ruggieri 1973; Massa et al. 2011). At the same time, it is isolated biogeographically and is characterized by numerous endemic species of flora and fauna (Pignatti 2011). Environmental adaptation and/or natural selection (climatic factors, bottleneck effect, genetic drift, gene flow, or random evolutionary processes caused by neutrality) have produced profound coevolutionary changes in all species in the region (Sineo et al. 2015). Homo sapiens was no exception and even before the intense cultural and biological contributions left by the Greek, Carthaginian, Roman, Byzantine, Islamic, and Norman/Swabian colonizers (Lauria et al. 2023), early Sicilians were subjected to and strongly influenced by the environmental conditions. These conditions quickly and significantly influenced the island’s gene pool and the human phenotype as early as Prehistory (D’Amore et al. 2009, 2010; Galland et al. 2019).
In recent years, the debate about the early human peopling of Sicily (and the related dispersal patterns) has almost exclusively focused on the archaeological evidence. The possibility for and degree of admixture caused by ancient migratory flows have been only investigated in limited anthropological studies conducted on a short time spans (D’Amore et al. 2009, 2010; Galland et al. 2019). The archaeological based settlement hypotheses assume an early colonization of Sicily’s northern coast during the Palaeolithic, initially driven by economic needs and the related subsistence strategies (Beretta et al. 1986; Beloch 1989; Di Salvo et al. 2008; Barucco et al. 2021). This was followed by a new colonization and partial substitution during the Mesolithic (Galland et al., 2019). The island was then significantly impacted by Neolithic and post-Neolithic demic movements (Fernandes et al. 2020) that determined the colonization of the hinterland and of the southeast coast during protohistoric periods. These movements were driven by cultural and social factors (Serratti 2000; Di Salvo et al. 2008) such as the foreign invasion of the island which was carried out by several populations (Siculi, Ausoni, Morgeti and Elimi – Holloway 2002).
Recent craniofacial morphometrics analyses (Lauria and Sineo 2023) that considered migratory flows and population influx have provided a more comprehensive approach to understanding settlement patterns. These analyses go beyond archaeologically based settlement hypotheses by merging previous archaeological evaluations and paleoclimatic studies with an anthropological approach.
This study expands the sample upon earlier morphometric work (D’Amore et al. 2009, 2010; Galland et al. 2019; Lauria and Sineo 2023) and provides an overview focusing the period from the Upper Palaeolithic to the Iron Age (14.500 B.C.E. – 900-800 B.C.E.). To reconstruct the biological history of the early human settlement in Sicily we analysed the shape variation (polarity and magnitude) of facial features in order to associate the changes in cranial morphology with population influx. Hypotheses were formulated by performing a Procrustes coordinates PCA to understand the separation between the specimens and the groups reasonably. Instead, PCA is an exploratory analysis we use its similarity with the discriminant analyses to draw our conclusions.
Materials and methods
For this study, human skulls from Sicilian Prehistory and Protohistory were considered. Approval to examine the skulls was issued by Department of Sicilian Cultural Heritage and the Gemmellaro Archaeological Museum. These skulls were divided into six periods (Table 1) based on the 14C cal B.P for SanTeodoro 1 (Sineo et al. 2002), Molara (Leighton 1999) and Oriente (Modi et al. 2022) and based on archaeological artifacts (industry and pottery) for SanTeodoro 2, Uzzo, Marcita and Polizzello (Leighton 1999) (Table 2). Sample selection was based on a detailed bibliographic review of previously published archaeological and historical works as well as previous anthropological and palaeoecological studies (Table 3). Additionally, the A. Salinas, Baglio Anselmi, and L. Bernabo’ Brea’ museum catalogues were consulted. A preliminary visual examination of the skeletal remains was carried out to assess their morphological completeness and to determine if they were suitable for inclusion in this study. To avoid any error related to approximation, broken, incomplete (skulls not in anatomical connection and/or lacking landmarks necessary for the GM study discussed below) and restored skulls were a priori excluded and not considered for the study.
| Main Sicilian Prehistoric and Protohistoric Periods: |
|---|
| B.C.E. Before Common Era |
| Prehistory |
| • Upper Paleolithic: 38.000–8.000 B.C.E. • Mesolithc: 8.000–6.000 B.C.E. • Neolithic: 6.000–4.000 B.C.E. • Eneolithic/Copper Age: 4.000–2.500 B.C.E. |
| Protohistory |
| • Bronze Age: 2.500–1.100 B.C.E. Early Bronze Age: 2.500–2.000 B.C.E. Middle Bronze Age: 2.000–1.500 B.C.E. Late Bronze Age: 1.500–1.100 B.C.E. • Iron Age: 1.100–700 B.C.E. |
| Site | Key | Specimens | 14C Dating cal B.P. | Periods |
|---|---|---|---|---|
| Cave of San Teodoro | ST | 2 | 14.500 | Upper-Paleolithic |
| Cave of Uzzo | Uz | 2 | – | Mesolithic |
| Cave of Molara | Mo | 1 | 8.600 | Mesolithic |
| Cave of Oriente | Or | 1 | 10.544 | Mesolithic |
| Ragusa | Ra | 3 | – | Eneolithic(Copper Age) |
| Marcita | Ma | 4 | – | Bronze Age |
| Polizzello | Po | 2 | – | Iron Age |
| Archaeological and historical works | Anthropological and palaeoecological studies |
|---|---|
| Belvedere et al. 2017 | Becker 1995–2000 |
| Bonfiglio et al. 2001 | Castellana & Mallegni 1986 |
| Borgognini et al. 1985–1993 | Di Salvo 1991 |
| Borgognini & Repetto 1986 | Di Salvo et al. 1998–2007–2012 |
| Chilardi & Galdi 2012 | Galland et al. 2019 |
| Conte et al. 2017 | Garilli et al. 2020 |
| Costantini 2014 | Incarbona et al. 2010a–2010b |
| De Miro 1988 | Mannino et al. 2017 |
| Hodos 2018 | Messina et al. 2008 |
| La Rocca 2011 | Miccichè et al. 2018 |
| Panvini et al. 2020 | Schimmenti & Di Salvo 1997 |
| Tusa 1994 | Sineo et al. 2002–2005 |
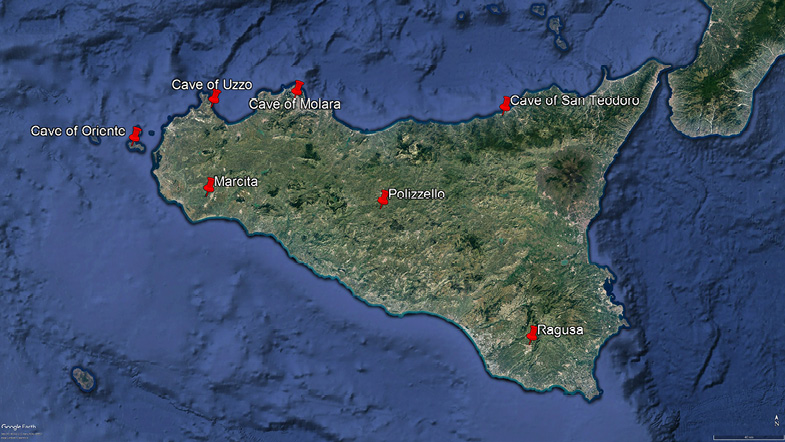
In total, 16 adult (Ubeleker 1989; Scheuer and Black 2000; Buikstra and Ubelaker 1994) human skulls (Table 4) from eight different settlements were selected (Figure 1, Table 2). The spatial bias in the map (Figure 1) is mainly due to a lack of specimens from the southeastern part of the island, which saw scarce colonization (lack of settlements) of Homo before the historical periods. Despite the small number of specimens and settlements, the sample effectively represents the inhabited regions of Sicily before the historical conization.
| Site | Key | Label | Sex |
|---|---|---|---|
| Cave of San Teodoro | ST1 | ST1 | Female |
| no data | ST2 | ST2 | Male |
| Cave of Uzzo | Uz1 | Uz4a | Female |
| no data | Uz2 | Uz5 | Male |
| Cave of Molara | Mo | Mo2 | Male |
| Cave of Oriente | Or | OrB | Male |
| Ragusa | Ra1 | RaT2S2 | Male |
| no data | Ra2 | RAT2S1 | Male |
| no data | Ra3 | RaT2S3 | Female |
| Marcita | Ma1 | MaTC8 | Male |
| no data | Ma2 | MaTC18 | Male |
| no data | Ma3 | MaTC20 | Female |
| no data | Ma4 | MaTC16 | Female |
| Polizzello | Po1 | No label | Female |
| no data | Po2 | No label | Male |
With the aim of associating the diachronic changes in cranial morphology (Relethford 2002, 2004a, b; Roseman 2004; Taubadel and Lycett 2008; von Cramon-Taubadel 2008; 2009a; 2009b; 2009c; 2014; Betti et al. 2009; Smith 2009; 2011; von Cramon-Taubadel and Weaver 2009; Matsumura et al. 2018; Manthey and Ousley 2020; Klingenberg 2022) with population migrations, we performed a 3D geometric morphometrics (GM) comparative analysis. This study followed the approaches proposed by several authors (Bruner and Manzi 2004; Bruner 2007; D’Amore et al. 2009, 2010; Baab et al. 2010; Smith 2011; Fredline et al. 2012; Matusmura et al. 2018, 2022; Galland et al. 2019; Hubbe et al. 2020; Lauria and Sineo 2023; von Cramon-Taubadel and Lycett, von Cramon-Taubadel 2008, 2009a, 2011, 2014, 2017; Grine 2023a, 2023b; Gunz and Freidline 2023, Ribot et al 2023). Sex was estimated by evaluating the skulls’ morphological characters according to Acsàdi-Nemeskèri (1970) as reported in Walrath (2004) and Minozzi and Canci (2015). The 3D models from Grotta di SanTeodoro, Grotta della Molara, and Grotta D’Oriente were acquired using computed tomography (CT) while the specimens from Grotta dell’Uzzo, Ragusa, Partanna, Marcita, and Polizzello were obtained through photogrammetric reconstruction following the protocol proposed by Lauria et al. (2022). The method consists of a Structure-from-Motion (SfM) photogrammetry that uses a single camera to capture chromatic details and reconstruct shape. A series of photos were taken forming circles around the target that will completely be covered and reconstructed by allying the photos and building a cloud of point before the polygonization of the model in the end.
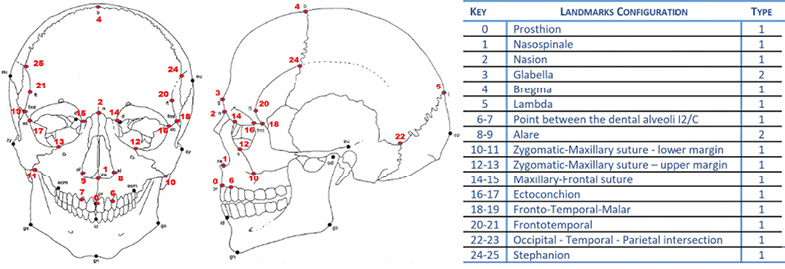
Photogrammetric and CT models were scaled with the software used to build the models (Metashape 1.5.1 for photogrammetry and Slicer 5.2.2 for CT) and after exported in PLY format. The GM analyses were based on a configuration of 26 Landmarks (Buikstra and Ubelaker 1994) (Figure 2) positioned on the suture boundaries (Landmarks Type 1) and on the anthropometric points (Landmarks Type 2) (Bookstein 1991). Using the software “Landmark3.6” (Wiley et al. 2007), 78 Raw Coordinates (RCs) were acquired for each specimen and were then analysed using the software programs “MorphoJ 2.0” (Klingenberg 2011) and “PAST 2.0” (Hammer and Harper 2001).
MorphoJ was employed to subject the RCs (exported from “Landmark”) to a Generalized Procrustes Analysis (GPA) (Dryden and Mardia 2016) to remove the effects of translation and rotation and to standardize each specimen to unit centroid size (Gower and Payne 1975, Rohlf and Slice 1990, Goodall 1991). The resulting Procrustes Fitted Coordinates (PFCs) were visualized using shape change graphs, including Lollipop and Wireframe Graphs, which allow three-dimensional forms to be visualized in two dimensions (Hammer and Harper 2008; Klingenberg 2013). These graphs illustrated the shape changes from a starting shape (the mean shape in the sample) to the target shape (the most extreme of the specimens) (Klingenberg 2013). These graphs were used to capture shape variation (direction and magnitude) across time (Harvati et. 2007, 2010, Bruner and Ripani 2008, Baab et al. 2010, Galland et al. 2016, 2019, Lauria and Sineo 2023). To highlight the positions of the specimens and groups within the sample (Hammer and Harper 2008), the RCs (exported from MorphoJ) were procrustized again in PAST to perform a Principal Component Analysis (PCA) based on a Covariance Matrix (represents the change of each variable relative to the others, including itself). The PCA separately analysed the PFCs of the single specimens, the average PFCs of each site, and the average PFCs of period.
Always considering that PCA is an exploratory analysis that allows for the formulation of a hypothesis based on the visualization (Le Maître and Mitteroecker 2019) and that discriminant analyses were not applicable (MANOVA and CVA are possible only when the variables are less than the specimens, Bronstain et al. 2006, Hammer and Harper 2008), we used the similarity between PCA and other discriminant analyses (Lauria and Sineo 2023) to reasonably understand the separation between the specimens and the groups. PAST was also employed to create a cluster analysis (CA) (UPGMA cluster procedure and Euclidean distance matrix) (Saitou and Nei 1987), leaving the software to recognize the outgroup.
Finally, on the PFCs, were conducted two “one-way ANOVA tests”, the Levene’s test for homogeneity of variance (Levene 1960; Derrick et al. 2018) and the more robust Shapiro-Wilk test (Shapiro and Wilk 1965) to evaluate the null hypothesis (H0) of equal multivariate means between the groups.
Results
Eigenvalue and percentage of variance
The plots of the PFCs (Single Specimens: Figure 3a; Averages of Each Site: Figure 3b; Averages of Each Period: Figure 3c) show that only the first two Principal Components (PCs) are significant. In detail, for the single specimens, the % of variance is 38,317% and 26,252% and the eigenvalue stands at 37,4197 and 26,252%, respectively. For the average of each site the % of variance is 38,188% and 26,717% while the eigenvalue are 33,4632 and 23,4111. Finally considering the averages of each period the values are 43,531% and 24,462% for the % of variance and 28,2558 and 15,8780 for the eigenvalue. In general, both the eigenvalues and the percent variance decrease gradually, converging toward similar values (Figure 3d-f), with a slightly more marked trend between the first two PCs (Figure 3). This suggests greater variations between the Paleolithic and the Mesolithic components (hunter-gatherers of the Stone Age) compared to the farmers-shepherds of the Metal Ages.
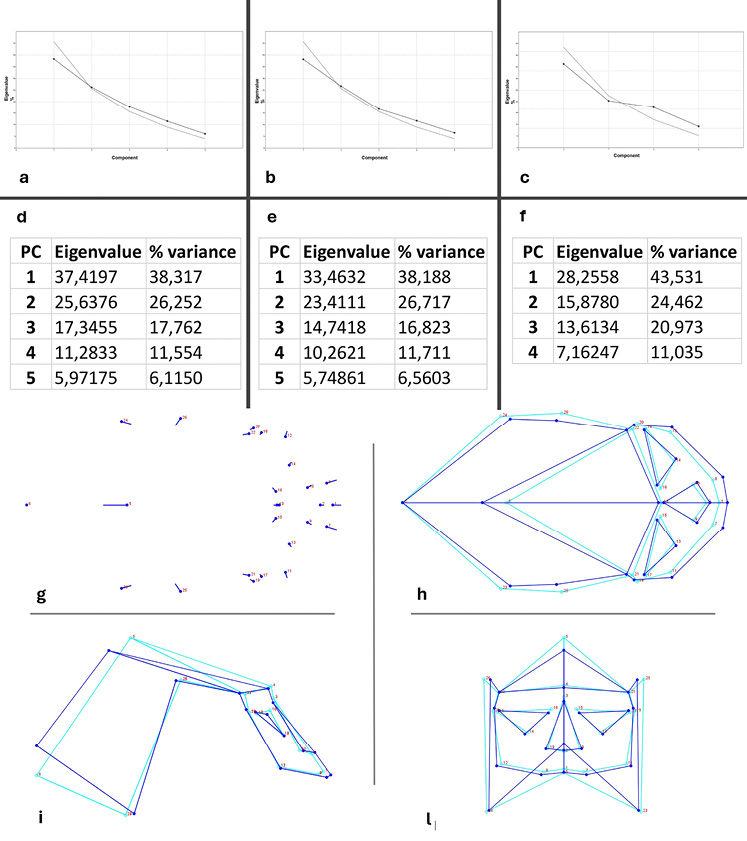
Shape variation
Considering that sample size and sample composition (number of specimens and sexual dimorphism) can influence the analysis, the shape variations show that the most notable changes are located on the neurocranium and the lower face, while minor changes affected the upper face (landmarks 4 and 19–22), the nose, and the orbits (Figure 3g). The frontal bone generally increased in size, becoming more elongated and lower (Figure 3h-i), while the width remained almost unchanged (Figure 3l). The parietal bones slightly reduced in size, showing decreases in length, the height of the posterior portion (Figure 3i), and in the width of the superior part (Figure 3l). The nose became slightly elongated (Figure 3g), however its size and position remained almost unchanged (Figure 3i-l). The orbits showed minimal changes in size and position (Figure 3l), only moving slightly downward.
Principal component analyses and cluster analyses
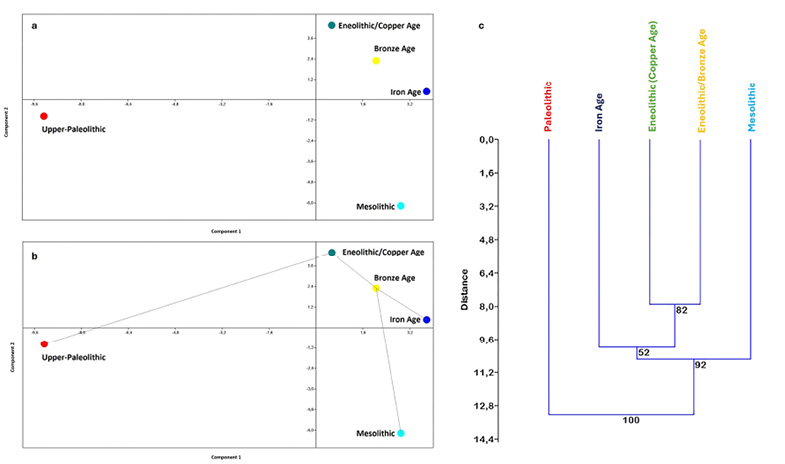
The PCA performed on PFCs of the single specimens (Figure 4a-b) shows the two Upper Paleolithic specimens from San Teodoro (Incarbona et al. 2010a) lying exactly on the negative axes of PC1, separated from the other specimens. In contrast, all the Mesolithic hunter-gatherer specimens from Molara, Uzzo, and Oriente are located on the positive axes of PC1. The Uzzo specimens are positioned on the negative side of the PC2 axes and those from Molara and Oriente are on the positive side of the PC2 axes. Notably, the Oriente specimens appear separated from both the Molara ones and all other individuals. In general, the hunter-gatherer specimens from the Sicilian Stone Age occupy an inhomogeneous morphospace surrounding the specimens representing the farmers-shepherds of the Sicilian Metal Ages. The farmers-shepherds of Ragusa (Copper Age/Eneolithic), Marcita (Bronze Age), and Polizzello (Iron Age) are all grouped in a homogenous morphospace near the centre of PCs axes (Figure 4a-b).
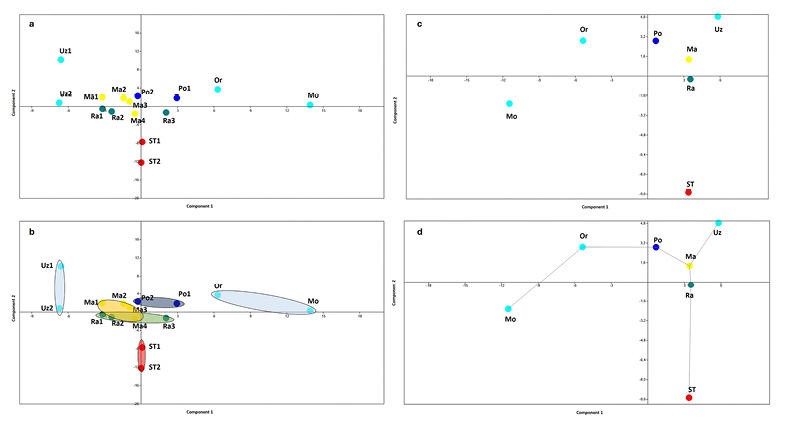
Further PCA analyses, based on the averages of the PFCs (average of each site: Figure 4c-d; average of each period: Figure 5a), reinforce the previous findings; the Paleo-Mesolithic sites of San Teodoro, Molara, Uzzo, and Oriente each occupy different quadrants in the PCA, surrounding the Copper, Bronze, and Iron Age sites of Ragusa, Marcita, and Polizzello, respectively (Figure 4c-d). Among these last three sites (all positioned on the positive PC1 axis), it should be noted that the Iron Age site of Polizzello appears slightly distant from the sites of Ragusa and Marcita, which are close to each other but separated by the PC2 axis (Figure 4c-d). These results also show a significant distance between the Paleolithic and Mesolithic periods (both hunter-gatherers, separated by the PC1 axis; Figure 5a), which in turn are far from the Copper, Bronze, and Iron Ages (all farmers-shepherds), grouped together in the upper right quadrant (positive PC1 and PC2 axes) close to each other but each in their own position (Figure 5a). The CA results (Figure 5b), based on the averages of the PFCs for each period, confirm the PCA findings. San Teodoro is automatically recognized as the outgroup, while the Paleolithic group retains some affinities with the Mesolithic specimens, which cluster separately from both the Paleolithic and the more recent farmers-shepherds from the Metal Ages. Additionally, the Prehistoric Eneolithic/Copper Age and the Protohistoric Bronze Age are still clustered together and, in turn, separated from the Iron Age group, which is chronologically closest to the most recent historical periods.
One-way ANOVA
The Levene’s test for homogeneity of variance to evaluate the H0 returned in all the case p (same) value < 0.001 (in a significance level of α=0.05), in detail PFCs specimens p (same)=3,569E-31, PFCs sites p (same)=1,915E-10 and PFCs periods p (same)=9,223E-12 that rejects the H0. In the same way the Shapiro-Wilk test returns PFCs specimens p (same)=6,067E-19, PFCs sites p (same)=1,6E-12 and PFCs periods p (same)=1,016E-7 that also in this case rejects the H0.
Discussion
Overall, the gradual decrease in eigenvalues and the percent variance suggests a slow progressive increase in variability (Figure 3) caused by primitive low-density migrations. These migratory events of the Paleolithic and Mesolithic (Stone Age hunter-gatherers) exhibit spatial clustering distinct from the later Metal Age farmer-shepherds.
The shape variations across prehistory and protohistory (Figure 3g) show a general trend of dolicefalization of the neurocranium (becomes more elongated and narrower). In contrast, the lower face became more elongated but wider, while the nose and orbits largely maintained their size and position. This pattern suggests phenotypic and genotypic changes due to neutral forces such as environmental factors and the stochastic forces that generally evolve in a neutral manner (Smith 2011). These acted in parallel with the population influx caused by migratory flows from the content (Betti et al. 2009). The scatterplots generated by the PCAs (Figures 4–5) and by the CA (Figure 5) show a decrease in the biological distance from the Paleolithic to the Iron Age, with short distances between the Sicilian Metal Age groups. Specifically, the hunter-gatherer migratory flows of the Stone Ages (Paleolithic and Mesolithic) were always characterized by sporadic, low-density migrations that produced cyclical and discontinuous occupations of the island. From the Eneolithic/Copper Age onward, there was a slow increase in both frequency and density of migratory flows, continuing through the Bronze and Iron Ages. Although these demic migrations produced some population discontinuity, the concentric arrangement indicates a limited but constant degree of admixture among the mentioned groups. This implies no significant morphometric variation between Prehistory and Protohistory populations. Finally, these dynamics produced a not negligible allometry supported by the one-way ANOVA tests, that both returned p (same) values < 0.001 that rejects the H0.
Although some of our landmarks (such as glabella or inion) involves sexually dimorphic cranial areas males and females have not influenced the scatterplots. Indeed, no differences were observed between groups from all sites. Male and female individuals from the same site are often very close to each other (Galland et al. 2019). When evaluating the variation between human groups arriving to a localized geographical region (like an island), it is important to consider that the genetic pool is often stressed by genetic drift phenomena such as the bottleneck and founder effect (Manica et a. 2007). In addition to these stochastic forces, adaptive changes (such as the masticatory-inducted phenotype) are in parallel impacted by cultural variations with the same plasticity but with a slow degree of diversification (Harvati and Weaver 2006). In particular, patterns of the cranial vault and the upper face are evolving largely neutrally (Smith 2011). Nevertheless, the large differentiation of facial shapes during the centuries could not only be explained by adaptive changes but also by the arrival of new genetic components (Betti et al. 2009).
Conclusion
According to the paleoclimatic data (during the last glacial peak, Sicily was characterized by a steppe or semi-steppe environment and extremely low rainfall values) (Incarbona et al. 2010a, b; Sadori et al. 2008) a stable occupation by Homo sapiens of the island was possible not before the Upper–Paleolithic. Right in that period early migratory flows arrived from the continent on the northwest coast and continued exclusively along the northern coastline, moving east to west in a counterclockwise direction. PCA analyses show that, during the Sicilian Stone Age, the hunter-gatherer’s colonizers cyclically occupied the land establishing settlements in the proximity of caves, close to the coastlines. Moreover, the scatterplot displays that only during the transition to a mobile-forager/semi-sedentary ecology during protohistoric period allowed a gradual increase in the frequency and density of migratory flows. Although the degree of admixture was limited, semi-migratory farmers-shepherds began abandoning caves for small villages. This shift marked the start of the colonization of the southeast cost and hinterlands, a process that was only completed in the first historical periods with Greek and Punic colonization (700 B.C.E.).
References
Acsádi GY, Nemeskéri, J. 1970. History of human life span. Budapest.
Baab KL, Freidline SE, Wang SL, Hanson T. 2010. Relationship of cranial robusticity to cranial form, geography and climate in Homo sapiens. Am J Phys Anthropol 141(1):97–115. https://doi.org/10.1002/ajpa.21120
Barucco D, Battaglia G, Battiato S, Bazan G, Bettelli M, Biondi G, Vita G. 2021. La Sicilia preistorica: Dinamiche interne e relazioni esterne. Atti del convegno internazionale di studi: Catania-Siracusa 7–9 ottobre.
Becker MJ. 1995. Skeletal studies of Sicilian populations. A survey. J Accord Res Inst 6:83–117.
Becker MJ. 2000. Skeletal studies of the people of Sicily. An update on research into human remains from archaeological contexts. Int J Anthropol 15:191–239. https://doi.org/10.1007/bf02445134
Beloch J. 1889. La popolazione antica della Sicilia. Libreria Internazionale.
Belvedere O, Burgi A, Bordonaro G, Forgia V. 2017. Baucina (Pa)–Monte Falcone 2014 Indagini nella necropoli. In FOLDandR; FastiOnLine Documents and Research. Roma, Italy.
Beretta M, Mazzetti P, Frosina G, Schilirò G, Russo A, Russo G, Barrai I. 1986. Population structure of eastern Sicily. Hum hered 36(6):379–387. https://doi.org/10.1159/000153662
Betti L, Balloux F, Amos W, Hanihara T, Manica A. 2009. Distance from Africa, not climate, explains within-population phenotypic diversity in humans. Proc Roy Soc B. 276: 809–814. https://doi.org/10.1098/rspb.2008.1563
Bonfiglio L, Marra AC, Masini F, Petruso D. 2001. Depositi a vertebrati e ambienti costieri pleistocenici della Sicilia e della Calabria meridionale. Biogeogr J Integr Biogeogr 22:29–43. https://doi.org/10.21426/B6110178
Bookstein FL. 1991. Landmark methods for forms without landmarks. Morphometrics of group differences in outline shape. In Medical Image Analysis; Elsevier: Amsterdam, The Netherlands, Volume 1:225–243. https://doi.org/10.1016/S1361-8415(97)85012-8
Borgognini SM, Elena R. 1985. Dietary patterns in the Mesolithic samples from Uzzo and Molara caves (Sicily). The evidence of teeth. J Hum Evol 14:241–254. https://doi.org/10.1016/S0047-2484(85)80065-8
Borgognini SM, Repetto TE. 1986. Skeletal indicators of subsistence patterns and activity régime in the Mesolithic sample from Grotta dell’Uzzo (Trapani, Sicily): A case study. Hum Evol 1:331–351. https://doi.org/10.1007/BF02436707
Borgognini SM, Canci A, Piperno M, Repetto E. 1993. Dati archeologici e antropologici sulle sepolture mesolitiche della Grotta dell’Uzzo (Trapani). Bullettino di Paletnologia Italiana; Istituto Poligrafico e Zecca Dello Stato: Roma, Italy 84:85–179.
Bosch R, Real J, Tinto A, Zozaya EL, Castell C. 2010. Home‐ranges and patterns of spatial use in territorial Bonelli’s Eagles Aquila fasciata. Ibis. 152(1):105–117. https://doi.org/10.1111/j.1474-919X.2009.00985.x
Bruner E. 2007. Cranial shape and size variation in human evolution: Structural and functional perspectives. In Child’s Nervous System; Springer: Boston, MA, USA 23:1357–1365. https://doi.org/10.1007/s00381-007-0434-2
Bruner E, Manzi G. 2004. Variability in facial size and shape among North and East African human populations. Ital J Zool 71:51–56. https://doi.org/10.1080/11250000409356550
Bruner E, and Ripani M. 2008. A quantitative and descriptive approach to morphological variation of the endocranial base in modern humans. Am J Phys Anthropol 137(1), 30–40. https://doi.org/10.1002/ajpa.20837
Buikstra JE, Ubelaker DH. 1994. Standards for Data Collection from Human Skeletal Remains. Arkansas Archaeological Survey: Fayetteville, AR, USA 71–73.
Castellana G, Mallegni F. 1986: The Prehistoric Settlement of Piano Vento in the Territory of Palma di Montechiaro (Agrigento, Italy). Arch. Per L’antropologia E La Etnol. 116:61–80.
Chilardi S, Galdi A. 2012. Il Neolitico siciliano tra caccia e raccolta, tradizione e innovazione: la fauna delle UUSS 1–16 del fossato di Contrada Stretto/Partanna (TP). Atti della XLI Riunione Scientifica dell’Istituto Italiano di Preistoria e Protostoria Dai Ciclopi agli Ecisti. Società e territorio nella Sicilia preistorica e protostorica (San Cipirrello, Palermo 2006) 1121–1125.
Conte L, Ingoglia AK, Martìn AML, Riolo L. 2007. Il culto dell’acqua e la civiltà dei fossati nel territorio di Partanna (Trapani). Annali dell’Università degli Studi di Ferrara 1824–2707.
Costantini L. 2014. Plant exploitation at Grotta dell’Uzzo, Sicily: New evidence for the transition from Mesolithic to Neolithic subsistence in southern Europe. In Foraging and Farming; Routledge: London, UK 197–206.
von Cramon-Taubadel N, Lycett SJ. 2008. Human cranial variation fits iterative founder effect model with African origin. Am J Phys Anthropol 136:108–113. https://doi.org/10.1002/ajpa.20775
von Cramon-Taubadel N, Weaver TD. 2009a. Insights from a quantitative genetic approach to human morphological evolution. Evol Anthropol 18:237–240. https://doi.org/10.1002/evan.20233
von Cramon-Taubadel N. 2009b. Congruence of individual cranial bone morphology and neutral molecular affinity patterns in modern humans. Am J Phys Anthropol 140:205–215. https://doi.org/10.1002/ajpa.21041
von Cramon-Taubadel N. 2009c. Revisiting the homoiology hypothesis: The impact of phenotypic plasticity on the reconstruction of human population history from craniometric data. J Hum Evol 57:179–190. https://doi.org/10.1016/j.jhevol.2009.05.009
von Cramon-Taubadel N. 2011. The relative efficacy of functional and developmental cranial modules for reconstructing global human population history. Am J Phys Anthropol 146:83–93. https://doi.org/10.1002/ajpa.21550
von Cramon-Taubadel N. 2014. Evolutionary insights into global patterns of human cranial diversity: Population history, climatic and dietary effects. J Anthropol Sci 92:43–77. https://doi.org/10.4436/JASS.91010
von Cramon-Taubadel N, Strauss A, Hubbe M. 2017. Evolutionary population history of early Paleoamerican cranial morphology. Sci Adv 3:e1602289. https://doi.org/10.1126/sciadv.1602289
D’Amore G, Di Marco S, Tartarelli G, Bigazzi R, Sineo L. 2009. Late Pleistocene human evolution in Sicily: Comparative morphometric analysis of Grotta di San Teodoro craniofacial remains. J Hum Evol 56.537–550. https://doi.org/10.1016/j.jhevol.2009.02.002
D’Amore G, Di Marco S, Di Salvo R, Messina A, Sineo L. 2010. Early human peopling of Sicily. Evidence from the Mesolithic skeletal remains from Grotta d’Oriente. Ann Hum Biol 37(3), 403–426. https://doi.org/10.3109/03014461003712947
De Miro E. 1988. Polizzello, centro della Sicania. In QuadMess; Università Degli Studi di Messina. Istituto di Archeologia: Messina, Italy 3:25–41.
Derrick B, Ruck A, Toher D, White P. 2018. Tests for equality of variances between two samples which contain both paired observations and independent observations. J App Quant Meth 13(2):36–47.
Di Salvo R. 1991. Tre Resti Cranici da Marcita. Archivio Per L’Antropologia e la Etnologia. Società Italiana per l’Antropologia e la Etnologia: Firenze, Italy, Volume 122:251–258.
Di Salvo R, Germanà F, Tusa S. 1998. Uomini e Culture Della Sicilia Preistorica; Gaia Editrice: Milano, Italy.
Di Salvo R, Schimmenti V, Mannino M, Caramelli D, Lalueza-Fow C., Messina A, ... and Sineo L. 2007. Ecologia, morfometria e genetica dei reperti paleomesolitici di Grotta D’Oriente (Favignana, TP). Atti XVII Congresso degli Antropologi Italiani Mediterraneo, crocevia di popoli e culture. Processi microevolutivi delle popolazioni umane, 52–53.
Di Salvo R, Mannino G, Mannino MA, Schimmenti V, Sineo L. 2012. Le sepolture della Grotta d’Oriente (Favignana). Le sepolture della Grotta d’Oriente (Favignana), 341–351.
Dryden IL, Mardia KV. 2016. Statistical Shape Analysis. With Applications in R. John Wiley and Sons: Hoboken, NJ, USA 995. https://doi.org/10.1002/9781119072492
Freidline SE. 2012. Evolution of the face in Mid Pleistocene Homo-3D surface analysis of ontogeny, allometry and evolution. PhD thesis. City Univ. New York.
Galland M, Van Gerven DP, Cramon-Taubadel V, Pinhasi R. 2016. 11,000 years of craniofacial and mandibular variation in Lower Nubia. Sci Rep 6(1).1–10. https://doi.org/10.1038/srep31040
Galland M, D’Amore G, Friess M, Miccichè R, Pinhasi R, Sparacello VS, Sineo L. 2019. Morphological variability of Upper Paleolithic and Mesolithic skulls from Sicily. J Antropol Sci 97:151–172. https://doi.org/10.4436/JASS.97009
Garilli V, Vita G, La Parola V, Vraca MP, Giarrusso R, Rosina P, Sineo L. 2020. First evidence of Pleistocene ochre production from bacteriogenic iron oxides. A case study of the Upper Palaeolithic site at the San Teodoro Cave (Sicily, Italy). J Archaeol Sci 123. https://doi.org/10.1016/j.jas.2020.105221
Goodall C. 1991. Procrustes methods in the statistical analysis of shape. J R Stat Soc Ser B Methodol 53:285–321. https://doi.org/10.1111/j.2517-6161.1991.tb01825.x
Gower JC, Payne RW. 1975. A comparison of different criteria for selecting binary tests in diagnostic keys. In Biometrika; Oxford University Press: Oxford, UK 62:665–672. https://doi.org/10.2307/2335526
Grine FE. 2023a. Description and comparative morphology of the Hofmeyr skull. Hofmeyr A Late Pleistocene human skull from South Africa. Cham: Springer International Publishing: 71–118. https://doi.org/10.1007/978-3-031-07426-4_6
Grine FE 2023b. Introduction. The Fossil Record of Homo sapiens in Africa–Morphological Variability in the Late Quaternary and the Significance of the Hofmeyr Skull. Hofmeyr A Late Pleistocene Human Skull from South Africa. Cham: Springer International Publishing:1–5.
Gunz P, Freidline SE. 2023. Cranial form of the Hofmeyr skull: Comparative 3D geometric morphometrics. Hofmeyr A Late Pleistocene human skull from South Africa. Cham: Springer International Publishing: 143–150.
Hammer Ø, Harper DA, 2008. Paleontological data analysis. John Wiley and Sons.
Hammer Ø, Harper DA, Ryan PD. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica; Coquina Press: Minneapolis, MN, USA, 4:9.
Harvati K, Weaver TD. 2006. Human cranial anatomy and the differential preservation of population history and climate signatures. Anat Rec 288(12): 1225–1233. https://doi.org/10.1002/ar.a.20395
Harvati K, Gunz P, Grigorescu D. 2007. Cioclovina (Romania): affinities of an early modern European. J Hum Evol 53(6):32–746. https://doi.org/10.1016/j.jhevol.2007.09.009
Harvati K, Hublin JJ, Gunz P. 2010. Evolution of middle-late Pleistocene human cranio-facial form: A 3-D approach. J Hum Evol 59(5):445–464. https://doi.org/10.1016/j.jhevol.2010.06.005
Hodos T. 2010. Globalization and Colonization: A View from Iron Age Sicily. J Mediterr Archaeol 23:81–106.
Holloway RR. 2002. The archaeology of ancient Sicily. Routledge.
Hubbe M, Terrazas Mata A, Herrera B, Benavente Sanvicente ME, González González A, Rojas Sandoval C, Von Cramon-Taubadel, N. 2020. Morphological variation of the early human remains from Quintana Roo, Yucatán Peninsula, Mexico: Contributions to the discussions about the settlement of the Americas. PloS one, 15(1):e0227444. https://doi.org/10.1371/journal.pone.0227444
Incarbona A, Zarcone G, Agate M, Bonomo S, Stefano E, Masini F, Sineo L. 2010. A multidisciplinary approach to reveal the Sicily Climate and Environment over the last 20,000 years. In Open Geosciences; De Gruyter: Berlin, Germany 2(40).
Incarbona A, Agate M, Arisco G, Bonomo S, Buccheri G, Di Patti C, Di Stefano E, Greco A, Madonia G, Masini F, et al. 2010. Ambiente e clima della Sicilia durante gli ultimi 20 mila anni. Alp Mediterr Quat 23:21–36.
Klingenberg CP. 2011. MorphoJ: An integrated Software Package for Geometric Morphometrics. Mol Ecol Resour 11:353–357. https://doi.org/10.1111/j.1755-0998.2010.02924.x
Klingenberg CP. 2013.Cranial integration and modularity: Insights into evolution and development from morphometric data. In Hystrix Ital J. Mammal; La Sapienza University: Roma, Italy 24(10).
Klingenberg CP. 2022. Methods for studying allometry in geometric morphometrics: a comparison of performance. Evol Ecol 36:439–470 https://doi.org/10.1007/s10682-022-10170-z
La Rocca P. 2011. Variabilità Cranimetrica e Distanze Biologiche tra Popolazioni Preistoriche ed Antiche Della Sicilia. PhD. Thesis, Universita’degli Studi di Catania, Catania, Italy.
Lauria G, Sineo L, Ficarra S. 2022. A detailed method for creating digital 3D models of human crania: An example of close-range photogrammetry based on the use of Structure-from-Motion (SfM) in virtual anthropology. Archaeol Anthropol Sci 14:42 https://doi.org/10.1007/s12520-022-01502-9
Lauria G, Sineo L. 2023. Human Peopling and Population Dynamics in Sicily: Preliminary Analysis of the Craniofacial Morphometric Variation from the Paleolithic to the Contemporary Age. Heritage 6:1187–1208. https://doi.org/10.3390/heritage6020066
Leighton R. 1999. Sicily before history: an archaeological survey from the Palaeolithic to the Iron Age. Cornell University Press.
Le Maître A, Mitteroecker P. 2019. Multivariate comparison of variance in R. In Methods in Ecology and Evolution; British Ecological Society: London, UK, Volume 10:1380–1392. https://doi.org/10.1111/2041-210X.13253
Levene H. 1960. Robust tests for equality of variances. In Ingram Olkin; Harold Hotelling; et al. (eds.). Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. Stanford University Press: 278–292.
Manica A, Amos W, Balloux F, Hanihara T. 2007. The effect of ancient population bottlenecks on human phenotypic variation. Nature, 448(7151): 346–348. https://doi.org/10.1038/nature05951
Mannino MA, Thomas KD, Leng MJ, Piperno M, Tusa S, Tagliacozzo A. 2017. Marine resources in the Mesolithic and Neolithic at the Grotta dell’Uzzo (Sicily): Evidence from isotope analyses of marine shells. Archaeometry 49:117–133. https://doi.org/10.1111/j.1475-4754.2007.00291.x
Manthe L, Ousley SD. 2020 Geometric morphometrics. In: Statistics and probability in forensic anthropology. Academic Press: 289–298.
Massa B, Sbordoni V, Vigna Taglianti A. 2011. La Biogeografia della Sicilia. Proceeeding of the XXXVII Congresso della Societa’Italiana di Biogeografia, Catania, Italy, 1–7 ottobre 2018. Biogeogr J Integr Biogeogr 30:685–694.
Matsumura H, Shinoda KI, Shimanjuntak T, Oktaviana AA, Noerwidi S, Sofian HO, Adachi N. 2018. Cranio-morphometric and aDNA corroboration of the Austronesian dispersal model in ancient Island Southeast Asia: Support from Gua Harimau, Indonesia. PLoS ONE 13:e0198689. https://doi.org/10.1371/journal.pone.0198689
Matsumura H, Tanijiri T, Kouchi M, Hanihara T, Friess M, Moiseyev V, Miyahara K. 2022. Global patterns of the cranial form of modern human populations described by analysis of a 3D surface homologous model. Sci Rep 12(1):13826. https://doi.org/10.1038/s41598-022-15883-3
Messina A, Sineo L, Schimmenti V, Di Salvo R. 2008. Cribra Orbitalia and Enamel Hypoplasia of the Iron Age (IX–VII centuries BC) Human Group of Polizzello (Sicily). J Palaeopathol 20:53–65.
Miccichè R, Carotenuto G, Sìneo L. 2018. The utility of 3D medical imaging techniques for obtaining a reliable differential diagnosis of metastatic cancer in an Iron Age skull. Int J Paleopathol 21:41–46. https://doi.org/10.1016/j.ijpp.2017.03.006
Minozzi S, Canci A. 2015. Archeologia dei resti umani: dallo scavo al laboratorio. Carocci.
Modi A, Vizzari, MT, Catalano G, Boscolo Agostini R, Vai S, Lari M, Ghirotto S. 2022. Genetic structure and differentiation from early bronze age in the mediterranean island of sicily: Insights from ancient mitochondrial genomes. Frontiers in Genetics, 13, 945227.
Panvini R, Scerra S, Terranova G. 2020. Un insediamento eneolitico nel territorio di Ragusa. Vivere all’ombra del Vulcano, 253.
Pignatti S. 2011. La flora della Sicilia come chiave di lettura per la fitogeografia mediterranea: Una visione autobiografica. Biogeogr J Integr Biogeogr 30:71–94
Relethford JH. 2002. Apportionment of global human genetic diversity based on craniometrics and skin color. Am J Phys Anthropol 118:393–398. https://doi.org/10.1002/ajpa.10079
Relethford JH. 2004a. Global patterns of isolation by distance based on genetic and morphological data. Hum Biol 76:499–513. https://dx.doi.org/10.1353/hub.2004.0060
Relethford JH. 2004b. Boas and beyond: migration and craniometric variation. Am J Hum Biol 16:379–386. https://doi.org/10.1002/ajhb.20045
Ribot I, Ghalem Y, Crevecoeur I. (2023). The position of the Hofmeyr skull within Late Pleistocene and Holocene African regional diversity: 2D and 3D morphometric analyses. In Hofmeyr: A Late Pleistocene human skull from South Africa. Cham: Springer International Publishing 119–141.
Rohlf FJ, Slice D. 1990. Extensions of the Procrustes method for the optimal superimposition of landmarks. Systemc Biol 39:40–59. https://doi.org/10.2307/2992207
Ruggieri G. 1973. Due parole sulla paleogeografia delle isole minori a Ovest ea Nord della Sicilia. Biogeogr J Integr Biogeogr 3:5–12.
Saitou N, Nei M. 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454
Scheuer L, Black S. 2000. Developmental juvenile osteology. Elsevier Ltd., London.
Schimmenti V, Di Salvo R. 1997. Progetto del Laboratorio Osteologico per l’archivio informatizzato.
Serrati J. 2000. Sicily from pre-Greek times to the fourth century. Sicily from Aeneas to Augustus. New Approaches in Archaeology and History, 9–14. https://doi.org/10.3366/edinburgh/9780748613670.003.0002
Shapiro SS, Wilk MB. 1965. An analysis of variance test for normality (complete samples). Biometrika 52(3–4): 591–611.
Sineo L, Bigazzi R, D’Amore G, Tartarelli G, Di Patti C, Berzero A, Caramella Crespi V. 2002 I resti umani della Grotta di S. Teodoro (Messina): Datazione assoluta con il metodo della spettrometria gamma diretta (U/Pa). In Antropo; University of the Basque Country: Bilbao, Spain 2:9–16.
Sineo L, Petruso D, Forgia V, Messina A, D’Amore G. 2015: Human peopling of Sicily during quaternary. In Quaternary Period; AcademyPublish.org: London, UK 25–67.
Smith HF. 2009. Which cranial regions reflect molecular distances reliably in humans? Evidence from three-dimensional morphology. Am J Hum Biol 21:36–47. https://doi.org/10.1002/ajhb.20805
Smith HF. 2011. The role of genetic drift in shaping modern human cranial evolution: a test using microevolutionary modeling. Int J Evol Biol 2011(1):145262. https://doi.org/10.4061/2011/145262
Surdi G. 2011. I Micromammiferi Tardo Quaternari della Sicilia e del Sud della Penisola Italiana.
Tusa S, Valente I. 1994. ’La ricerca archeologica in contrada Stretto-Partanna: il fossato/trincea neolitico. La Preistoria del Basso Belice e della Sicilia meridionale nel quadro della preistoria siciliana e mediterranea. Società Siciliana di Storia Patria. Palermo 177–195.
Walrath DE, Turner P, Bruzek J. 2004. Reliability test of the visual assessment of cranial traits for sex determination. Am J Phys Anthropol 125(2):132–137. https://doi.org/10.1002/ajpa.10373
Wiley DF, Armenta N, Alcantara DA, Ghosh D, Kil YJ, Delson E, Harcour-Smith, WH, Rohlf FJ, Hamann B, St John K. 2007. Landmark 3.6. In Institute of Data Analysis and Visualization (IDAV); University of California: Davis, CA, USA.
Final information
Acknowledgements
None.
Funding
None.
Conflicts of interest
The authors declare that they have no conflict of interest and no competing interests.
Ethics statement
The interpretations reported in the present study are based on the analysis of skeletal findings obtained through excavations and authorized by institutional permits.
Statement of contributions from authors
Conceptualization and Investigation: G.L. and L.S.; Methodology and Software: G.L.; Formal Analysis and Data Curation: G.L.; Data Interpretation, Writing and Editing: G.L. and L.S.; Supervision, Funding acquisition and Project administration: L.S.
Corresponding author
Gabriele Lauria, Department of Biological, Chemical and Pharmaceutical Sciences and Technologies (STEBICEF), University of Palermo, Via Archirafi 18, 90123 Palermo, Italy; e-mail: gabriele.lauria03@community.unipa.it