 https://orcid.org/0000-0003-3649-3989
https://orcid.org/0000-0003-3649-3989
Introduction
Suture closure ages are notoriously variable in humans (Meindl and Lovejoy 1985; Buikstra and Ubelaker 1994; Hershkovitz et al. 1997). When the premature ossification of suture mesenchyme occurs in prenatal or early postnatal development, the cranium continues to grow, but largely within those neurocranial regions unconstrained by premature suture closure, resulting in an abnormal vault morphology (Marsh et al. 1991; Cohen 2005; DeLeon and Richtsmeier 2009; Ursitti et al. 2011; Kajdic et al. 2018; Eisová et al. 2021). Partial or complete ossification of the sagittal mesenchyme during early cranial development results in a visibly narrow head shape or scaphocephaly, often including elevation of the fused suture (Weber et al. 2008; Ursitti et al. 2011; Soficaru et al. 2018).
Scaphocephaly is often the result of sagittal synostosis although the converse is not necessarily true (Cohen 2005; Padmalayam et al. 2013; Groza et al. 2014). Sagittal synostosis is detectable by the third trimester and is the most common form of craniosynostosis, affecting approximately 1 in 5,000 births (Constantine et al. 2000), and is 3.5 times as common in males compared to females (Cohen 2005; Ursitti et al. 2011; Kajdic et al. 2018). Partial synostosis of the sagittal suture occurs in about a third of cases among infants around one year (Boyajian et al. 2020). As infants age, the partial closure of the sagittal sutures often proceeds to full fusion.
Scaphocephaly has been found in several Neolithic contexts across Europe. The earliest of these include the Eneolithic of Spain and the Neolithic cultures of central Europe (Table 1). The paucity of individuals noted to exhibit scaphocephaly point to the lack of evidence of head binding in comparison to other prehistoric contexts, such as the Neolithic Near East and Pre-Columbian Mesoamerican, Hopi and Andean cultures (Antón 1989; Meiklejohn et al. 1992; Kohn et al. 1994; Gerszten et al. 1998). Here we report a case of scaphocephaly, possibly from sagittal synostosis or partial fusion (bridging), found in an early adolescent cranium from the Neolithic cemetery of Modřice. Scaphocephaly in Modřice 3871 is coupled with other craniofacial pathologies, including facial asymmetry, the presence of wormian bones, cribra orbitalia and porotic hyperostosis of the temporal bone.
| Site | Location | Identifier | Dating | Sex | Age in years |
| Kónya | Hungary | SNR 247/1 | Linear Pottery Culture | ? | 6–7 |
| Kónya | Hungary | SNR 247/2 | Linear Pottery Culture | ? | 7 |
| West Kennet Barrowb | UK | EU 1.5.148 | Neolithic | F | 17–25 |
| Zengövárkonyc | Hungary | Grave Nr. 7 | Lengyel | F | 20–25 |
| Náměšť na Hanéd | Czechia | Feature Nr. 7 | Funnel Beaker | F | 30–50 |
| Cova del Palanquese | Spain | Skull Nr. 6 | Eneolithic | ? | ? |
aTóth et al. (2009); bCuthbert (2018); cSmrčka et al. (2018); dPankowská et al. (2010); eCampillo (1993)
Materials and Methods
Modřice cemetery
Modřice is located in the southern Moravia region of Czechia (Fig. 1). Extensive excavations recovered a rich prehistoric settlement and burial ground associated with the Funnel Beaker culture. The graves include skeletal remains, mostly without artifacts, and are radiocarbon dated to 3,800–3,600 years BCE (Šmíd et al. 2018, 2021a; Drtikolová Kaupová et al. 2023), corresponding to the late Neolithic or Early Eneolithic in Czech archaeology (Jarošová et al. 2022). The Funnel Beaker culture is associated with a further Neolithization of the incipient agricultural economy that rapidly spread into parts of northwest and northcentral Europe. The Funnel Beaker culture of Modřice was an agrarian community that included domesticated animals, wheat and vegetable cultivation as well as wild foods in the diet.
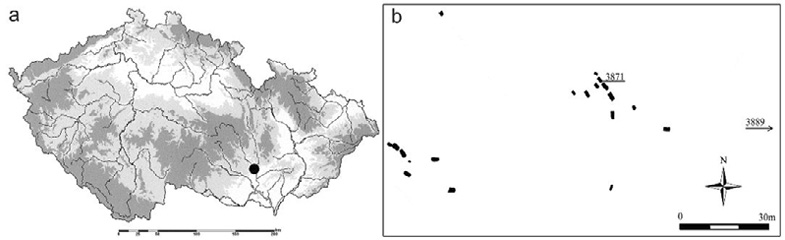
Fig. 1. Map of (a) Czechia showing the location of the Neolithic cemetery of Modřice indicated by the black circle; and (b) the cemetery showing the two main groups of graves and the position of Modřice 3871 vis-à-vis the other graves
At Modřice, there were 22 individuals buried in two main groups of single graves, each apparently comprising members of the same household (Šmíd et al. 2018, 2021a); two of the burials were in a settlement pit and no commingled graves were found. Individuals of both sexes were distributed across the two groups, about a half of whom were immature, including an early adolescent, Modřice 3871. In total there were five children 3–6 years-old, four older children 7–14 years, two adolescent females, six adult males and five adult females. Most of skeletons (including 3871) were deposited in an outstretched position, which was a common burial practice in this phase of the Funnel Beaker culture, although only recently recognized in Moravian archaeology (Šmíd et al. 2021b). The rectangular grave pit of 3871 was oriented NW-SE, and the burial was directly dated to 4880±33 years uncalBP (Drtikolová Kaupová et al. 2023). A subsequent Bronze Age pit located in the lower part of the legs disturbed the grave of Modřice 3871, and the bones were partially scattered, probably by animals (rodents). Some of the skeletal remains were partly fractured and many smaller bones of the hands and feet were missing (Fig. 2).
There are multiple paleopathologies found at Modřice, including spina bifida, spondylolysis and ankylosis of the lumbar vertebrae, entheses, clavicle and cervical asymmetry, healed and perimortem fractures, osteophytosis, osteoporosis, periosteal reactions and other skeletal abnormalities (Šmíd et al. 2021a, Table V). The dental health noted at the Modřice cemetery was rather poor with a relatively greater prevalence of caries compared to other Neolithic population from Moravia and East Bohemia (Jarošová et al. 2022).
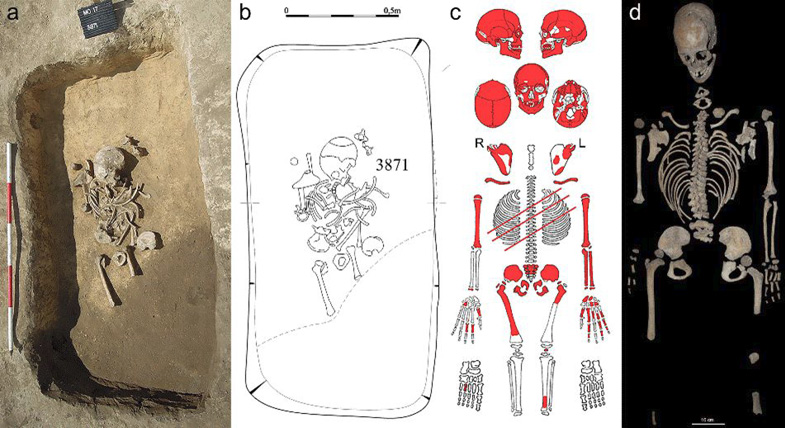
Fig. 2. Modřice 3871 (a) photograph and (b) drawing showing the skeleton in situ; (C) scheme of preservation, R = right, L = left; and (d) skeletal elements in anatomical position
Methods
Modřice 3871 was examined at the Moravian Museum in Brno, Czechia (Inv. nr. A25632) using macroscopic investigation to estimate the age and describe the preservation and morphology. Dental wear scores following Smith (1984) were used to characterize molar attrition to corroborate the age estimate. Radiographic images of Modřice 3871 were obtained at Fakultní nemocnice U sv. Anny v Brně (St. Anne’s University Hospital in Brno), and these were compared to the macroscopic examination of the cranial vault.
Results
Inventory
The grave of Modřice 3871 consists of a single incomplete early adolescent skeleton. The cortical surface is slightly eroded in places, and covered with calcareous debris. The bones are brownish-yellow in color (Fig. 2).
Cranial vault
The cranium of Modřice 3871 is relatively well preserved and partially reconstructed (Fig. 3). The cranial vault base is slightly deformed by the position of the basi-occipital bone, which is conjoined with the two lateral occipitals and slightly rotated to the left. The foramen magnum is nearly intact excepting a posterior margin with possible rodent gnaw marks.
The frontal and occipital bones are complete, whereas the parietal bones are reconstructed after recent fracturing in the area of the parietal tubercles. The damage is confined mostly along the parietal-occipital margin, and extends to the right occipitomastoid region. The right temporal bone is almost complete, whereas the left is damaged. The sphenoid body and greater wings are partly preserved.
The cranial vault is markedly expanded anteroposteriorly from a maximum cranial length (gonion to opistocranion = 185 mm) much greater than the maximum cranial breadth (euryon to euryon = 124 mm). The occipital extends posteriorly forming a relatively concave inferior contour in which the inferior squama abruptly tapers anteriorly forming a small occiput, with a sigmoid curvature extending further inferiorly to opisthion (Fig. 3).
Face and dentition
The relatively vertical frontal squama exhibits slight but palpable superciliary arch development. Much of the splanchocranium is missing, although the palate connects to the mid-face (Fig. 3). The posterior mid-face is partly damaged owing to the destruction of the inferior left and right palatine bones and the adjoining maxillary palatal border, while the mandible is complete.
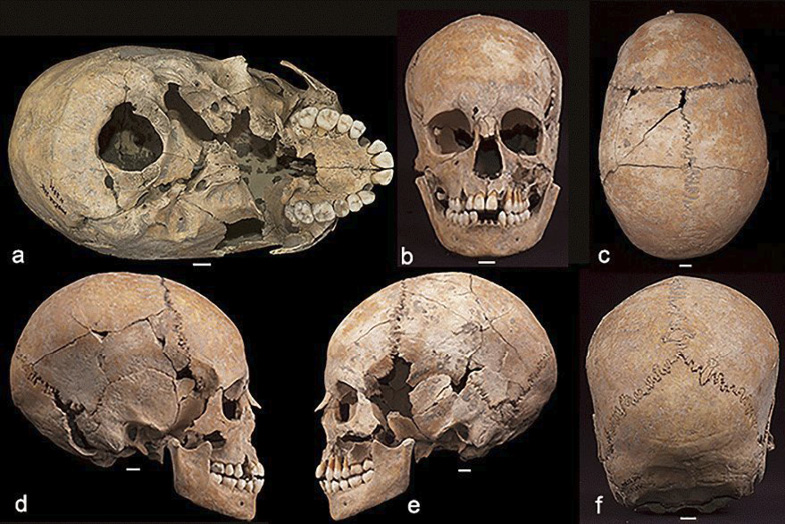
Fig. 3. Modřice 3871, (a) inferior view showing the elongated cranial vault; as well as (b) anterior; (C) superior; (d) lateral right; (e) lateral left; and (f) posterior planes; scale bars = 1 cm
The permanent dentition is mostly preserved in situ. On the maxilla, only the right canine and left I2 are missing postmortem, while the unerupted M3 is visible on both sides. On the mandible, only I1 on both sides and left I2 are lost postmortem.
Postcranial skeleton
The vertebrae ribs, scapulae and clavicle are well preserved. The humeri are also nearly complete and include separated proximal epiphyses without any fusion evident. From the right forearm, only the distal epiphysis of the radius is preserved. The forearm bones of the left are intact except for the proximal epiphyses. Five carpal bones, 5 metacarpals and 9 phalanges are preserved (Fig. 2).
The ischium and pubis are fused but are separated from the ilium. The right femur is largely preserved but misses the distal end of the shaft and epiphysis, although the greater and lesser trochanter and head epiphyses are present. There are gnaw marks on the diaphysis of the right femur. On the left femur, only the proximal third of the diaphysis is preserved, as is the distal diaphysis of the left tibia and the right fifth metatarsal (Fig. 2).
Age and sex estimation of Modřice 3871
Based on visual examination of the size of the cranium, the small and orthogonal face, and lack of coronal and lambdoial ossification (Fig. 3), the individual is aged as an early adolescent (Scheuer and Black 2000; Scheuer 2002). All of the permanent teeth have erupted with the exception of the third molars, which are deeply embedded within the crypts, although the crowns are fully formed and the growth of roots has commenced. The permanent teeth, and in particular M2 and M2 show minimal attrition or Stage 2, whereas M1 is consistent with Stage 3 (Smith 1984).
The lack of fusion of epiphyses of the preserved appendicular skeleton provide additional corroboration of an immature age (Scheuer and Black 2000). These include the femoral head, the greater and lesser trochanters and distal tibial epiphysis, the fused ischiopubis and ilium articulation, the humeral head and distal trochlear end of the humerus, the proximal head and distal styloid processes of the radius and the proximal olecranon and distal styloid processes of the ulna (Fig. 2). Many of these epiphyses fuse during the late teens. However, the superior ischial and inferior pubic rami fuse at 6–9 years (Steele and Bramblett 1988), while the lesser trochanter of the femur and proximal radius commence fusion at 15–16 and 16 years respectively (Steele and Bramblett 1988), suggesting Modřice 3871 can be confidently aged between these extremes. The fact that the ischiopubic fusion site is still visible could suggests an individual not much older than 9 years. The triradiate cartilage joining the three innominate bones of Modřice 3871 is unfused (Fig. 2). The triratiate commences from multiple epiphyses, including a posterior one forming around 10–11 years, which eventually joins an anterior epiphysis around 12–14 years and these fuse with the epiphysis of the anterior inferior iliac spine, which forms at 14 years (Scheuer and Black 2004). The os coxa is completely ossified by 19 years (Scheuer and Black 2004). Modřice 3871 does not present any notable fusion of these triradiate epiphyses and lacks an anterior inferior iliac spine suggesting an age younger than 14 years. The unfused distal humerus which commences ossification at 14 years (Steele and Bramblett 1988) further corroborates an early adolescent attribution. There are no instances of the skeletal remains showing epiphyseal fusion that would be unusual for an early adolescent. The length of the long bones and state of fusion of all the epiphyses taken together indicate an age 10–13 years (Scheuer and Black 2000) while tooth eruption, including the slight wear on the fully erupted second molar, indicates an age closer to 14 years (Al Qhatani et al. 2010). The lack of sexually diagnostic features on the pelvis, cranium and mandible and the immature age of the individual prevented an estimation of sex membership.
Diagnosis
Although taphonomic processes have fractured and slightly distorted the cranium, the unusual elongated shape of the vault is unambiguous (Fig. 3). Weber et al. (2008) calculate a mean cranial length/breadth index for untreated scaphocephalic adult crania from historical and archaeological collections to be 63.4 (n = 18). The mean length/breadth index for adult crania without stenostosis is 78.9 (n = 40) (Weber et al. 2008). The length/breadth index for Modřice 3871 is 67.0, which is lower than then mean of 74.3 for the six complete adolescent and adult crania from Modřice (Šmíd et al. 2021a). The cranial index of the other adolescent from Modřice excavated from grave 3858 (female, 15–19 years) is 80.3.
The superior surface of the orbital wall exhibits cribra orbitalia (Fig. 4) indicative of nutritional stress. The superior ceiling of both orbits presents cribra orbitalia Grade 1–2 (Rivera and Lahr 2017) or Type 2–3 in the antero-intermediate sector following Stuart-Macadam (1991) (Fig. 4). Porosity is also evident in the area of porus accusticus externus.
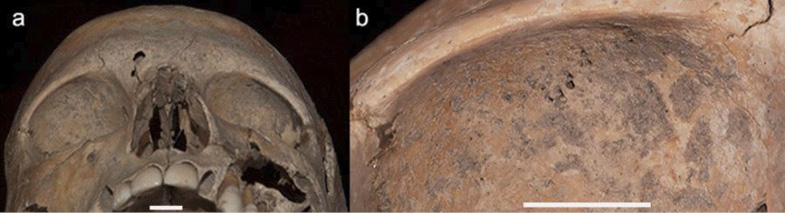
Fig. 4. Cribra orbital (a) viewed inferior on the orbital ceiling of the frontal bones; (b) the right orbital ceiling at a higher objective shows the lesions appear to be healing; both scale bars = 1 cm
In addition, the upper facial skeleton also appears slightly deformed. The right supraorbital rim is higher and more bulbous than the left in inferior view (Fig. 3 and Fig. 4). This difference is likely not the result of taphonomic processes since the bony table is mostly uninterrupted across the supraorbital margin. The right orbit is also shaped differently than the left. Whereas the left is relatively circular, the right orbit is wider, shorter, and more trapezoidal in shape (Fig. 4). There is also asymmetry in the partly preserved superior portions of the nasal bones in which the right is broader than the left.
Additional asymmetry exists in the superior shape of the cranial vault, such that the right lateral parietal arch bulges slightly laterally compared to the left (Fig. 3). In addition, the sagittal suture veers slightly to the left approaching the coronal suture about 10 to 15 degrees from the sagittal plane (Fig. 3). Although some of this distortion could possibly be from taphonomic factors, the slight but visible asymmetry at multiple locations on the upper face and vault, combined with mid-sagittal partial fusion could signal the presence of a syndromic synostosis involving multiple deformities (Ursitti et al. 2011; Kajdic et al. 2018). However, the coronal and lambdoidal sutures lack any evidence of closure, suggesting nonsyndromic synostosis (Giuffra et al. 2013) (Fig. 3).
The suture morphology is rugged and continuous, with the exception of an intervening wormian ossicle in the posterior sagittal suture (Fig. 3). Two additional wormian bones are located in the occipitomastoid sutures of the right and left sides (Fig. 3). Another double ancillary ossicle is found in the right lambdoidal suture (ossiculum suturae lambdoideae).
Radiography of Modřice 3871 corroborates the mid-sagittal partial closure endocranially (Fig. 5). Like the ectocranial surface, the endocranium features an unfused coronal, anterior sagittal and lambdoidal suture. The mid- and posterior sagittal suture areas are represented by a faint line that demarcates the fusion site and the lack of two independent parietal bones in Modřice 3871 (Fig. 5). The radiological and macroscopic analyses confirm that the mid-sagittal suture exhibits partial fusion endocranially and ectocranially, while the coronal and lambdoidal are open. Rather than the classic definition of sagittal synostosis (White 1996), it appear that the formation of bone spicules that cross the suture, or bridging, could be present (Fig. 5). The ectocranial surface presents a suture partly filled with matrix, suggesting an opening sufficient to collect debris (Fig. 5). Therefore, a diagnosis of bone bridging rather than synostosis is advanced to characterize the partial fusion of the sagittal suture.
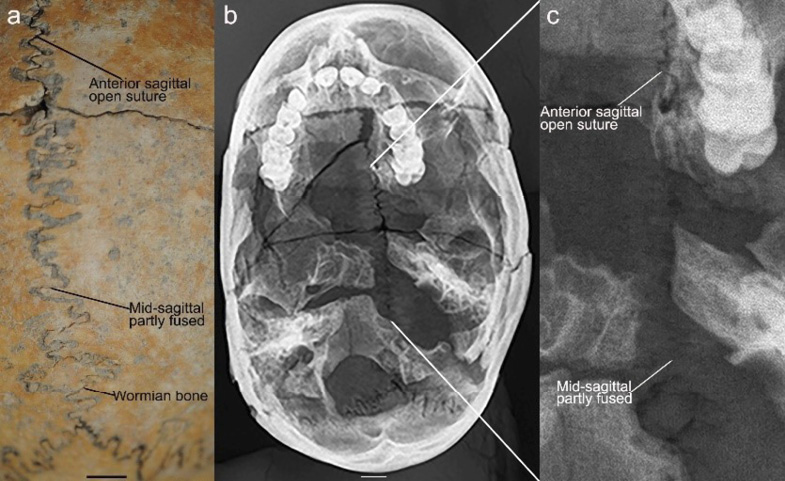
Fig. 5. Modřice 3871 cranial vault (a) superior view, showing the mid-sagittal suture ectocranially and a sagittal wormian bone; (b) radiograph of the cranium; and (C) enlarged area on the radiograph showing the mid-sagittal suture; scale bars = 1 cm
It is possible that Modřice 3871 suffered from Klippel-Feil syndrome given the asymmetry of the face and bridging of the mid-sagittal suture (Herrerín et al. 2022). Basilar impression is also associated with Klippel-Feil syndrome. The basilar region of Modřice 3871 is slightly displaced inferiorly. Yet Modřice 3871 lacks the concavity of the posterior occipital squama, sphenoid and petrous portion of the temporal surrounding the foramen magnum that characterizes basilar impression (Chamberlain 1939; Herrerín et al. 2022; Fig. 3). In addition, the cervical vertebrae do not show ankylosis, severe osteophytosis or any notable ossification (Fig. 2); the sagittal suture is not ossified along its entire length; and no large parietal foramina are present (Herrerín et al. 2022). Although craniofacial asymmetry and sagittal suture bridging exist, Klippel-Feil syndrome, which is not associated with the formation of wormian ossicles, is an unlikely diagnosis for Modřice 3871.
Discussion
Scaphocephaly is often the result of untreated sagittal synostosis, which is most common in the posterior and middle portion of the suture (Marsh et al. 1991; Cohen 2005; Ursitti et al. 2011). Constriction of the fetal neurocranium, including the base, during the third trimester does seem to correlate with sagittal synostosis, which may explain a greater prevalence in males who have larger skulls, even during gestation (White 1996). Constriction or mechanical stress results in a close alignment of cranial ossification centers during the sensitive periods of prenatal dura development that could influence the expression of sagittal synostosis (DeLeon and Richtsmeier 2009; Padmalayum et al. 2013).
Sagittal synostosis is one of the most common human craniofacial anomalies, and is usually apparent to external observers by the end of the first year (Gracia et al. 2009). By the second year of life, most cases of sagittal synostosis exhibit complete fusion of the sagittal suture (Kohn et al. 1994), although early closure of a sagittal suture does not necessarily result in scaphocephaly (Cohen 2005; Padmalayam et al. 2013; Groza et al. 2014). The degree of cranial elongation would depend on the timing of the closure.
Prehistory of scaphocephaly
Scaphocephaly has been noted since antiquity (Padmalayum et al. 2013), and was present at least as early as the Middle Pleistocene (Garcia et al. 2009). Modřice 3871 presents one of the most ancient examples of scaphocephaly although there are others. Three from Hungary are earlier. Two of these are from the latter half of the 6th millennium BCE (Tóth et al. 2009), and another is dated to 4,500–4,300 BCE (Smrčka et al. 2018). At Náměšť na Hané, a cranium dated to a slightly earlier time period but also from a Funnel Beaker culture settlement in Moravia, exhibits sagittal synostosis and is associated with a cranial index of 55 (Pankowská et al. 2010). Relative to the female from Náměšť na Hané, Modřice 3871 may have presented mild scaphocephaly that probably arose from postnatal bridging of the sagittal suture.
Although head binding causes significant closure to the sagittal suture (White 1996), only one individual, Modřice 3871, out of both cemetery plots (Fig. 1) exhibits this elongated cranial form. Head binding was widely practiced in the Americas to denote status and identity (Antón et al. 1992; Meiklejohn et al. 1992; White 1996). Eurasian head binding has also been documented although more rarely and is often associated with cultures characterized by stricter hierarchies and greater contact among diverse peoples than would be typical for a small-scale Neolithic society like Modřice (Meiklejohn et al. 1992; Fernandes et al. 2019).
Wormian bones
Both tensile and compressive forces act to prematurely fuse the suture in affected individuals (White 1996). These forces also tend to be implicated in the formation of wormian bones (White 1996; Sanchez-Lara et al. 2007). Their presence is consistent with other studies noting wormian bones in individuals exhibiting premature sagittal fusion (White 1996). Sagittal and other synostoses as well as wormian bones, have been considered by some to be adaptive responses to prenatal constraints on the developing cranium (Moss 1959; White 1996; Pedersen and Antón 1998; Cohen 2005). Whether these ancillary ossicles are related to the presentation of partial fusion of the mid-sagittal suture is unknown. However, the presence of wormian bones do suggest some irregularities of sutural development during early embryogenesis.
However, one of the wormian bones of Modřice 3871 is found in the sagittal suture which is rare (Fig. 5). Most wormian bones occur in the lambdoidal suture or at lambda and less often in the occipitomastoid (Safak et al. 2020) and coronal sutures (Bellary et al. 2013). Some studies report an absence of sagittal sutural bones (Safak et al. 2020) whereas others demonstrate its presence albeit in small numbers (Vijay et al. 2017). Pryles and Khan (1979) suggest wormian bones to be a marker of central nervous system abnormality, craniosynostosis and other anomalies although its presence in 61% of individuals in one study (Vijay et al. 2017) and 59% in another (Cirpan et al. 2015) may indicate a lack of concordance between sutural ossicles and other disorders. However, the presence of several wormian bones may signal the existence of other anomalies (Cremin et al. 1982). This is the case for Modřice 3871 in which 5+ are found at multiple locations. Given the bridging of the mid-sagittal suture (Fig. 5), the presence of a sagittal wormian bone offers greater support for linking these anomalies to dural mechanical stress during maturation, either pre- or postnatally, that adversely affected cranial development. The placement of the sagittal wormian bone observed in Modřice 3871, approximately 2 cm superior to lambda, also approximates the position of the third fontanelle, which is associated with other birth defects (Chemke and Robinson 1969). It is possible that the sagittal wormian bone formed to compensate for a third fontanelle, although it remains unknown.
Cribra orbitalia and nutrition
The nutritional stress indicated by the presence of cribra orbitalia is also noteworthy. In addition to its manifestation on Modřice 3871, cribra orbitalia affected another 4 individuals from the cemetery, and nearly all are children (4 of 5). Altogether, cribra orbitalia was present in 38.5% of available individuals (5 of 13) indicating nutritional and/or disease stress broadly affected this Neolithic farming community, particularly its youngest members (Šmíd et al. 2021a). Other indices of nutritional stress include porosity of the temporal and asymmetry of the face. Furthermore, wormian bones were present in 8 out of 15 individuals (53.3 %) (Šmíd et al. 2021a), suggesting a possible genetic or behavioral influence on cranial suture formation in the community. Although the presentation of skeletal abnormalities at this cemetery is widespread (Šmíd et al. 2021a), the combination of paleopathologies in Modřice 3871 is unique.
The analysis of stable isotopes of individuals from Modřice shows that this community did not differ from other Funnel Beaker populations in the region, whose diet was of a terrestrial origin with a dominant C3-plant component supplemented with animal products (Drtikolová Kaupová et al. 2023). The isotope values for Modřice 3871 (δ15N = 9.5, δ13C = -19.8) fall close to the average of the Modřice population (δ15N = 9.5; δ13C = -19.6). Cribra orbitalia could have been the result of any number of processes, acting alone or in combination, including nutritional stress, anemia, infections and parasites, all of which are connected to a Neolithic agrarian lifestyle. The lack of agreement between older dental and younger epiphyseal age estimates may also point to nutritional stress.
Asymmetry of the basicranium
In Modřice 3871, the basicranium is titled to the left (Fig. 2). Although the possibility remains that taphonomic processes distorted the basicranium, it is unlikely given the corroborating evidence of asymmetry in the facial skeleton and superior cranial vault. Furthermore, the observed basicranial deformation may be associated with mid-sagittal bridging given the intertwined ontogenetic processes governing the formation of the vault and the base (Moss 1959; Antón 1989; DeLeon and Richtsmeier 2009; Ursitti et al. 2011). Asymmetry of the face and vault favoring the right side is consistent with the directionality of the foramen magnum and basion pointing to the smaller left side of the basicranium (Fig. 2). The combination of cranial asymmetry, scaphocephaly, multiple wormian bones, cribra orbitalia and porosity of the temporal are suggestive of multiple stressors during development, potentially including epigenetic and nutritional constraints.
Conclusion
The long narrow cranial shape, caused by endo- and ectocranial mid-sagittal bone bridging, indicates Modřice 3871 represents one of the oldest recorded instances of scaphocephaly in human history (Weber et al. 2008; Tóth et al. 2009; Pankowská et al. 2010; Smrčka et al. 2018). With respect to the lived experience, Modřice 3871 may have had an unusual head shape and slight facial asymmetry. The cribra orbitalia and porosity in the temporal bone indicates nutritional deficiencies or disease-related complications (Armelagos et al. 2014), which could have compromised the life expectancy of Modřice 3871. Nevertheless, Modřice 3871 was buried similarly to the others in the cemetery as has been reported for other prehistoric adolescents with visible congenital cranial anomalies (Pedersen and Antón 1998; Garcia and Santos 2020).
Acknowledgements
Research funding for Frank Williams was provided by the Department of Anthropology at Georgia State University. Zdeněk Tvrdý was funded through the institutional support of long-term conceptual development of research institutions provided by the Ministry of Culture (ref. MK000094862). We are grateful to Valerie DeLeon for valuable insights and the five anonymous reviewers whose comments improved the rigor of the paper.
Conflict of interest
The authors have no conflicts of interest to declare.
Authors’ contributions
The concept for the paper was developed by FLW and ZT in consultation with DP. The analysis, description and figures were prepared by FLW and ZT. The radiography was coordinated by ZT.

