Available online at: https://doi.org/10.18778/1898-6773.85.2.04
Independent researcher
University of Arizona, Tucson, Department of Psychology, AZ, USA
University of Arizona, Tucson, Department of Psychology, AZ, USA
Boston University, Boston, Department of Philosophy, MA, USA
Independent Researcher, Boston, MA, USA
ABSTRACT: The prediction that reduction of negative selection decreases group-level competitiveness, as reflected in increased individual-focused and diminished group-focused moral foundations, is tested. To measure this hypothesized shift in moral foundations, we conduct a culturomic analysis of the utilization frequencies of items sourced from the moral foundations item pool, tracked among Britannic populations from 1800 to 1999 using Google Ngram Viewer. The resultant higher-order factor, which tracks increasing individualizing values and decreasing binding values, is termed Asabiyyah (capturing social cohesion and collective purpose). Two predictors of this factor are examined: change in the strength of intergroup competition and change in levels of indicators of developmental instability. Both the strength of intergroup competition and levels of developmental instability associate with Asabiyyah. Rising developmental instability mediates the impact of inter-group competition, indicating that reduced between-group competition might have relaxed negative selection against mutations, which might reduce Asabiyyah via their effects on inter-genomic transactions. These results must be interpreted carefully, given the clear real-world evidence that explicit commitment to group-oriented values often features in harmful and maladaptive social and political ideologies of an extreme character.
KEY WORDS: Asabiyyah, Lexicographic data, Moral foundations, Multi-level selection
Social epistasis refers to intergenomic transactions that occur between at least two organisms, and which modify the gene expression of at least one of the involved organisms. Such social-epistatic changes of gene expression evidently can have phenotypic effects. For instance, in the work of Linksvayer (2007), an early user of the term “social epistasis,” evidence is reported that in “three species of closely related Temnothorax ants … adult worker size was determined by an interaction between the genotypes of developing brood and care-giving workers, i.e., intergenomic epistasis. Such intergenomic social epistasis provides a strong signature of coevolution between social partners” (p. 1). Evidence of social-epistatic effects has also been found in mice (see Bachmann et al. 2018; Cross 2019; Kalbassi et al. 2017). Among the most impressive findings concerns a mutation, specifically a gene deletion, in mice that is related to autistic-like behavior; it has been found that social association of mice who carry this mutation with mice who do not can lead to the latter exhibiting the autistic-like behavior of the former (Kalbassi et al. 2017).
Investigating the mechanism through which these behavioral changes in mice occur, Cross (2019) found evidence that social contact of mice that were carriers of this mutation with non-carriers changed RNA expression in the latter’s brain cells, which is consistent with a social-epistatic effect.
A relatively new line of primarily theoretical research into social epistasis in humans has concerned development of the social epistasis amplification model or SEAM (Woodley of Menie et al. 2017a). The SEAM was devised to offer a unified explanation of the apparent falling fitness and declining physical and mental health of Western populations from roughly the twentieth to twenty-first centuries (Woodley of Menie et al. 2017a). This model posits, first, that deleterious mutations have been accumulating in Western populations since the substantial relaxation of negative-selective pressure (i.e., selection that removes deleterious mutations) brought on by industrialization and many of its effects on social, technological, and economic development, especially improved sanitation and increases in wealth, which reduced burdens of infectious disease and general environmental harshness (for research on relaxed negative selection, see Kondrashov 2017; Lynch 2016; Rühli and Henneberg 2017). Second, it posits that the fitness costs of these accumulating deleterious mutations may be amplified via social epistasis, with certain deleterious mutations causing harmful changes in patterns of gene expression even in those who do not carry these variants, potentially reducing physical and mental health as well as reproductive success. A simulation indicates that this social-epistatic amplification of the fitness costs of a certain class of deleterious mutations, known as spiteful mutations because of their ability to externalize their fitness costs onto others via (in this instance) their social epistatic effect on the gene expression of other organisms in the same population[1], can theoretically cause very rapid decline in the fitness of entire populations (Woodley of Menie et al. 2017a).
One possibility is that differential expressions of moral foundations and associated systems[2], which Haidt (2007, see also Haidt 2012) defines broadly as “[i]nterlocking sets of values, practices, institutions, and evolved psychological mechanisms that work together to suppress or regulate selfishness and make social life possible”, may correspond to the relative strengths with which group- and individual-level selection have acted on populations over time, as well as their patterns of social epistasis. According to this hypothesis, greater expression of group oriented (or what Haidt (2012) calls binding) moral foundations, which are those prioritizing loyalty, sanctity, and respect for authority, reflects higher relative strength of group-level selection than greater expression of more individual-oriented (or what Haidt (2012) calls individualizing) moral foundations, which are those prioritizing fairness and avoidance of harm[3].
Hertler, Figueredo and Peñaherrera-Aguirre (2020), proposed that the interpretation of these chronometric factors as reflecting a culture’s orientation toward group- versus individual-level selection, resonates quite strongly with the ideas of medieval historian and sociologist Ibn Khaldun (1377), who believed that empires flourish when they are high in Asabiyyah. This concept describes a type of cohesive tribalism, and shares similarity with the concepts of esprit de corps and vigor, which denote a group’s legacy of toughness, grit, and resilience. Taken together, these traits contribute to exceptionally well-integrated groups which are thought to be more organized and capable in the face of conflict with rival groups. Ethnographic and historical evidence suggests that complex sociopolitical systems featuring ultrasocial mechanisms promoting within-group cooperation and proscribing social defection tend to outcompete simpler sociopolitical systems (Hertler et al. 2020). Asabiyyah as a prospective measure of groupishness conceptually overlaps with several subsets of terms that are characteristic of highly group-selected populations, including but not limited to élan, panache, and dash, as indicators of martial enthusiasm, comradery, loyalty, and compatriotism, as indicators of fraternal solidarity, and jingoism, nationalism, and patriotism, as indicators of national commitment and cohesiveness. The leixical basis of Haidt’s Binding higher-order moral foundation clearly also conceptually overlaps with Asabiyyah, with the production of words connoting loyalty, sanctity, and deference toward authority, serving as verbal-behavioral markers of orientation toward Asabiyyah. Conversely, when cultural emphasis is placed on concepts such as fairness and harm avoidance, reflecting an individualizing morality, this can be said to reflect an orientation away from Asabiyyah, as such values tend to be associated with personal flourishing rather than groupishness (Hertler et al. 2020). Ultimately, therefore, the term Asabiyyah simply denotes the distinctive observable quality of group-selected populations at the cultural level, and (based on arguments advanced in Hertler et al. 2020) this, in turn, might be reflected in a culture’s verbal behavior (e.g., in the generation of texts utilizing certain terms connoting a high binding and low individualising moral psychology).
Although complex polities initially benefit from the spoils of war, the influx of wealth and ease of living reduces a group’s level of Asabiyyah (Hertler et al. 2020; Khaldun 1377). Hertler and colleagues (2020) identified a stark macrohistorical decline in a lexicographic Asabiyyah factor across two centuries. According to the authors, GDP per Capita significantly reduced the level of Asabiyyah above and beyond any temporal autoregressive effects. This negative effect is expected since polities featuring greater macroeconomic growth and stability may allocate their available resources toward reducing morbidity and mortality rates. It follows then that such epidemiological transition should also be associated with relaxation of negative selective pressures facilitating the accumulation of deleterious mutations. Although consistent with SEAM, the authors of the latter study did not explore whether a reduction in between-group competition, an indicator of selective pressures, could positively influence (potential proxies for) mutation accumulation, which in turn may reduce Asabiyyah over time.
A prediction deriving from the SEAM is that Asabiyyah is likely to decline with time as a consequence of this relaxed negative selection leading to the accumulation of (in particular) spiteful mutations reducing the group-level cohesion of populations, and that this trend might be captured and measured lexicographically, as in Hertler et al. (2020), using changing cultural expressions associated with decreased binding morality, coupled with increased individualizing morality, marking the shift away from the sorts of values that are essential to the internal cohesiveness of groups (this being a key component of Asabiyyah).
There are also other factors that might influence a population’s level of Asabiyyah. Heightened expression of individualizing moral sentiment is very likely adaptive under low intergroup competition (peace), as reflected in the conditions that characterize Western late modernity (Hertler et al. 2020). This is likely because being more focused on the mitigation of harms and the promotion of fairness and personal flourishing, and a reduced emphasis on matters promoting group-level fitness, would be more beneficial to individuals under conditions of intergroup peace. This hypothesis is consistent with the evoked culture model of Tooby and Cosmides (1992), as flexibility in the development of moral foundations may constitute a kind of evolved plasticity which facilitates the adaptive calibration of behavior in response to various evolutionarily familiar environmental cues. Having gone through periods of both intergroup conflict and peace, human populations may have acquired behavioral and innate moral repertoires that adapt their members to both conditions, with such populations having become evolutionarily prepared for the expression of a range of moral sentiments that adaptively match the situation. Selection may also act via gene-culture coevolution to differentially promote the fitness of various moral genotypes under different regimes of group- and individual-level selection.
To compare these two models (the SEAM versus the evoked culture plus gene-culture coevolution model), in the current study Haidt’s moral foundations theory will be used to derive a lexicographic diachronic measure of Asabiyyah, as indicated by the decline in a latent common factor among the levels of both binding and (reverse scored) individualizing morality, measured via the utilization frequencies of words corresponding to Haidt’s moral foundations in the textual outputs of Britannic populations. It will then be determined whether a diachronic trend in measures of the strength of group-selection have direct effects on the level of this factor, or whether this is mediated by measures of increasing developmental instability (as a proxy for increasing mutation load). It is expected that the developmental instability factor should largely mediate the impact of the intergroup competition measure on the moral foundations factor. This is because decreased intergroup competitiveness likely relaxes negative selection via reductions in social conflict.
Data were collected for the following Britannic nations ranging from AD 1800 to 1999, essentially comprising the former British Empire and its various successor states: UK, USA, Canada, New Zealand, and Australia (Figueredo et al. 2019a).
The historical utilization of these specific classes of words was quantified via their relative frequencies of usage in English language texts across the 200 years spanning AD 1800–1999 through Google Ngram Viewer, an interactive textual corpus encompassing more than 5.9 million texts and 500 billion written words from AD 1500 to 2019 (Michel et al. 2011). The forward extent of our analysis is restricted to 1999, as the post-2000 corpus is known to be highly incomplete. This is consistent with other studies using this database (see: Greenfield 2013; Hills and Adelman 2015; Younes and Ulf-Dietrich 2019). Google Ngram Viewer has been used to track temporal trends in public sentiment (Figueredo et al. 2019a, 2019b; Greenfield 2013; Ladle et al. 2016; Michel et al. 2011), changes in expressions of religiosity (Younes and Ulf-Dietrich 2019), changes in population-level cognitive characteristics (as ascertained by the utilization frequencies of words with known item-level psychometric difficulties; see Roivainen 2014; Woodley of Menie et al. 2015 and historical estimates of word learnability; see Hills and Adelman 2015), shifts in lexicographically estimated life history characteristics (Woodley of Menie et al. 2019), and the temporal stability of cultural stereotypes (Del Giudice 2012). These applications of Ngram to the quantification and study of cultural trends are referred to as culturomics (Michel et al. 2011).
The lexical items connected with the moral foundations were retrieved from Graham, Haidt, and Nosek’s (2009) moral foundations dictionary: (1) harm, (2) fairness, (3) loyalty, (4) authority, and (5) purity (an abridged list of the words collected is presented in Table S1). The lexical items used in constructing each of these scales were psychometrically selected based on their possessing satisfactory part-whole correlations for each word to the corresponding aggregate scale score for each lexicographic scale. The best words were thus empirically selected from the initially larger item pool. This psychometric procedure for selecting items has the benefit of being the most straightforward approach to creating robust and internally consistent chronometric constructs, with the items exhibiting differential validity based on their degree of convergence with each of the five moral foundations.
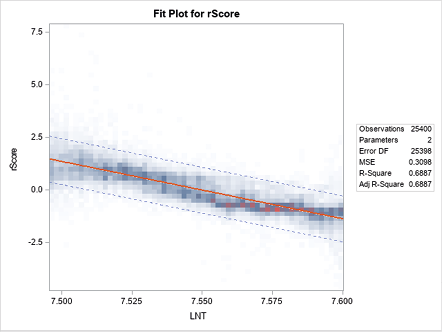
Unit-weighted common factor scales (Gorsuch 1983) were estimated as the means of the standardized scores for the lexicographic items on each scale (Figueredo et al. 2000). As per moral foundations theory, the five scales were aggregated into two lower-order factors: (1) binding and (2) individualizing. By reverse-scoring the individualizing factor, these two lower-order factors were further aggregated into a single higher-order factor—the Asabiyyah factor discussed in Hertler et al. (2020). The resultant chronometric factors, along with their Cronbach’s alpha values and unit-weighted factor loadings are presented in Table 1. Binding and individualizing exhibited a strong tendency toward negative correlation across time, indicating that these two trends were diverging from each other systematically. Therefore, there is a clear tendency for the rise in cultural expressions of morality emphasizing personal flourishing and self-actualization to occur at the expense of groupishness, deference to authority, and sanctity, which would be consistent with a decrease in Asabiyyah over time. This trend is graphed in Figure 1. It is important to note that the division of moral foundations into the categories of individualizing and binding does not reflect the explicit intent of the original authors to attribute these categories to individual versus group fitness; rather, this division more accurately describes alternate loci of moral values that both function to inhibit selfishness (Graham et al. 2011; Haidt 2008). The attribution of these diachronic changes to individual versus group selection was made by some of the current authors based on the application of multilevel selection theory (e.g., Hertler et al. 2020). Nevertheless, as discussed in the introduction, Haidt (2007, 2012) does speculatively attribute the evolution of binding values to cultural group selection, foreshadowing the present application. The logic expounded by Graham, Haidt, and Nosek (2009) is as follows: a society that takes a predominantly individualizing approach to suppressing selfishness will honor the rights and well-being of other individuals (care and fairness); in contrast, a society that takes a binding approach to suppressing selfishness will emphasize the imperative for individuals to conform to the needs of the group (loyalty, authority, sanctity/purity). If this reasoning is correct, then the implications for multilevel selection are quite manifest.
Fig. 1. Heat Map for Bivariate Linear Regression of Asabiyyah as a Natural Logarithmic Function of Time (AD 1800–1999).
Table 1 displays the psychometric results of these analyses. p-values here and later are based on two-tailed tests unless otherwise noted.
Table 1. Cronbach’s alphas and part-whole correlations (unit-weighted factor loadings) for the lexicographic scales and lower-order Moral Foundations factors of the higher-order Asabiyyah factor from AD1800–1999
| Cronbach’s Alpha | Unit-Weighted Factor Loading | |
| INDIVUALIZING | ||
| Harm Scale | 0.974 | 0.988* |
| Fairness Scale | 0.980 | 0.988* |
| BINDING | ||
| Loyalty Scale | 0.974 | 0.985* |
| Authority Scale | 0.991 | 0.995* |
| Purity Scale | 0.994 | 0.990* |
| ASABIYYAH | ||
| INDIVUALIZING | 0.988 | -0.987* |
| BINDING | 0.997 | 0.978* |
* p<.05
Following Woodley of Menie et al., (2017b), we also used the ten altruism words employed by Charles Darwin in The Descent of Man (1871) to describe changing levels of within-group altruism and between-groups competition in humans[4]. Diachronic part-whole correlations of the Darwin Descent of Man Altruism Words ranged from .29 to .92 (p<.05), with the overall factor scale accounting for 55% of the chronometric factor variance.
Population sizes were obtained from the Maddison Project database (Bolt at al. 2018), a repository curated by the Groningen Growth and Development Center. These were used to construct a corporate or group-level fitness measure by dividing the share of the Britannic populations by the rest of the world’s population at different points in time, yielding a relative measure of the success of their biocultural-group relative to the rest of the world’s population. The proportion of the world population was estimated based on the various demographic database compiled by Roser, Ritchie, and Ortiz-Ospina (2013, see also references therein).
Warfare mortality estimates (a fairly uncontroversial measure of inter-group competitiveness) were obtained from Sarkees and Wayman’s (2020) Correlates of War database, we excluded civil (within-state) conflicts, retaining only between-state conflicts. Any conflict involving one or more Britannic nation was retained. Mortality rates (expressed per capita, per 100,000) were estimated, controlling for population size. This was necessary, as population size could confound warfare intensity owing to the observation that larger populations will exhibit greater absolute death numbers.
Following Woodley of Menie et al. (2017b), a latent Intergroup Competition (IGC) Factor was constructed using the three convergent group-selection indicators: (1) Darwin’s Descent of Man Altruism Words; (2) Britannic corporate fitness; and (3) War Mortality per 100,000. The single lexicographic indicator used in the IGC factor model, the Darwin Descent of Man Altruism Words, converged well with the two biodemographic ones, the proportion of the world’s population and war mortality; part-whole temporal correlations of the IGC factor ranged from .42 to .66 (p <.05), with the overall factor scale cumulatively accounting for 27.5% of the chronometric factor variance.
This measure includes three convergent phenotypic measures that are believed to be associated (in part) with individual differences in burdens of deleterious mutations. The measures are percentage sinistrals (meaning those who are left-handed, sourced from McManus et al. 2010), craniofacial fluctuating asymmetry (sourced from Kimmerle and Jantz, 2006, with supplementary data from Woodley of Menie and Fernandes 2016), and body mass index (BMI; sourced from Komlos and Brabec 2010). The data were recovered from graphs in their respective publications using WebPlot Digitzer (Rohatgi 2017). Both sinistrality and fluctuating asymmetry have long been theorized to be indicators of developmental instability, and possibly also elevated mutational load (e.g., Markow 1992; van Valen 1962). The association between both the level and variance in BMI and deleterious mutations has only recently been evidenced however, with national-level indicators of relaxed negative selection functioning as substantial predictors of national differences in levels and variance of BMI (Budnik and Henneberg 2017), even when lifestyle covariates (e.g., calories consumed and levels of exercise) are controlled. Budnik and Henneberg (2017) have hypothesized that variation in BMI might be partly reflective of the action of deleterious variants that reduce the efficiency of metabolic processes, leading to either excessive body mass, or (in some cases) an inability to accumulate body mass.
In total, these variables are available for the years spanning 1825 to 1985. Sinistrality was sampled between the years 1835 to 1976, for a total of 99 measurement occasions. BMI was sampled between the years 1885 to 1985, for a total of 21 measurement occasions. Craniofacial fluctuating (specifically size) asymmetry was sampled between the years 1825 to 1985 for a total of 16 measurement occasions.
All variables were sourced from the population of the USA. The reason for focusing on the European-American samples in this instance is because the majority of the USA population for the majority of the set of years sampled here were of European descent. The incorporation of data on non-European-origin populations might therefore bias the sample characteristics in ways that are unrepresentative of the true time trends.
The use of phenotypes as proxies for tracking the underlying burden of deleterious mutations has been promoted in the absence of sufficiently high-resolution genomic sequencing and variant-calling protocols to detect the hypothesized increase in mutation accumulation that may have accompanied the reduction in opportunity for negative selection through mortality since industrialization (for discussion of this topic see Kondrashov 2017). It should furthermore be noted that factors independent of mutation accumulation may partly, and in some cases, mostly account for the time trends associated with these variables. For these variables to serve as useful proxies for mutation accumulation, it is only necessary that some of the temporal trends among them stem from relaxed negative selection however.
The results of the unit-weighted factor model (estimated using multivariate imputation; see Figueredo at al. 2000; McKnight at al. 2000) on the developmental instability factor are presented in Table 2. All factor loadings are statistically significant and high magnitude ranging in value from .68 to .99 (p < .05). The latent factor accounted for 75.5% of the chronometric variance.
Table 2. Part-whole correlations (unit-weighted factor loadings) for the Developmental Instability factor from AD1825-1985
| Unit-Weighted Factor Loading | |
| Developmental Instability | |
| Craniofacial Fluctuating Size Asymmetry | 0.68* |
| Body Mass Index | 0.91* |
| Sinistrality | 0.99* |
* p<.05
The following set of hypotheses are examined with reference to the data employed in the present study.
H1: Year will negatively predict the Intergroup Competition Factor.
H2: The Intergroup Competition Factor will negatively predict the Developmental Instability Factor.
H3: Year will positively predict the Developmental Instability Factor.
H4: The Developmental Instability Factor will negatively predict the Asabiyyah Factor.
H5: The Intergroup Competition Factor will positively predict the Asabiyyah Factor.
H6: Year will negatively predict the Asabiyyah Factor.
All univariate and multivariate analyses were performed using SAS 9.4 (SAS Institute Inc., 2015) and Unimult 2 (Gorsuch 2016). Using SAS PROC STANDARD and DATA, unit-weighted common factor scales (Gorsuch 1984) were estimated as the means of the standardized scores for all non-missing subscales on each factor (Figueredo et al. 2000). Using SAS PROC CORR, Cronbach’s alphas and the part-whole correlations of the subscales with the unit-weighted factor scales were also computed.
The lexicographic scales function as manifest variables for the purposes of longitudinally estimating multilevel models (MLMs). We estimated four nested MLMs in total, so as to determine the need for increasing parameterization as a function of testing alternative hypotheses. The four models are as follows: MLM1 was an unconditional Asabiyyah model, in which a single logarithmic slope and intercept were estimated for all lexicographic factors, scales, and items (words) over time. MLM2 involved the estimation of a separate intercept and logarithmic slope over time for each lexicographic factor. However, the same intercepts and logarithmic slopes were estimated for all within-factor scales and words over time. MLM3 involved the estimation of separate, lexicographic-scale-specific, logarithmic slopes and intercepts over time, but with each within-factor scale word having the same logarithmic slope and intercept over time. MLM4 involved the estimation of separate word-specific logarithmic slopes and intercepts over time. All MLMs were statistically controlled for the effects of the year of FirstUse recorded for each word in the analyses; this is an important control, as it has been found that older words tend to be better known to users of texts as a result of the lag between changes in spoken and written texts (Curzan 2009; Woodley of Menie et al. 2015); LNT is the natural logarithmic function of time.
All nested model comparisons are displayed in Table 3. Systematic -2RLL and AIC comparisons were performed by comparison among nested models. AIC and AIC weights were computed with the statistical package qpcR (Ritz and Spiess 2008) in R version 4.1.0. Each level of the aggregative hierarchy contains and accounts for specific variance components. Their estimation revealed that the majority of incremental model fit improvements were relatively trivial in magnitude, but nevertheless statistically significant (p<.05). When the four nested MLMs were compared in terms of the squared multiple correlations among them, it was found that they yielded basically the same results. Although statistically significant, the magnitudes of specific variances associated with each level of aggregation (ΔR2), were negligibly small, which contrasts sharply with the finding that the common factor variance associated with the highest-level of aggregation (unconditional Asabiyyah) was quite large (69%). It is worth noting that the model comparison identified MLM4, with Word and the Word*LNT interaction, as the best model based on its AIC weight (1.000).
Table 3. Fit Indices for Nested Multilevel Models (MLMs) for Haidt Moral Foundation Dictionary Factors, Scales, and Words from AD1800–1999
| Multilevel Model | MLM1: FirstUse + LNT |
MLM2: + Factor + Factor*LNT |
MLM3: + Scale + Scale*LNT |
MLM4: + Word + Word*LNT |
| AIC | 42323.8 | 42313.8 | 42307.1 | 41700.1 |
| AIC | 623.70 | 613.70 | 607.00 | 0.00 |
| AIC weight | 0.000 | 0.000 | 0.000 | 1.000 |
| -2RLL | 42313.8 | 42299.8 | 42281.1 | 41186.1 |
| Δχ2= | 14.0* | 18.7* | 1095.0* | |
| R2 | 0.68869 | 0.68886 | 0.68909 | 0.70221 |
| ΔR2= | 0.00017* | 0.00023* | 0.01312* | |
| NDF | 2 | 4 | 10 | 253 |
| ΔNDF= | 2 | 6 | 243 | |
* p<.05
Only the model parameters associated with the unconditional Asabiyyah level (MLM1) were retained, as the extra model parameters added by all the lower levels of aggregation (MLM2, MLM3, and MLM4) in the moral foundations dictionary factors, scales, and words only increased the proportion of variance explained from 69% to 70%. The logarithmic slope of this unitary higher-order Asabiyyah factor over time was negative and statistically significant: r = -.83, F(1,25398) = 56190.9, p<.0001. No significant heterogeneous serially autoregressive effects were identified (ARH1 = 0), and the effect of year of FirstUse (of each word) was statistically nonsignificant (p>.05).
We constructed a sequential canonical cascade model from the following system of three ordered hierarchical multiple regressions:
The purpose of a sequential canonical cascade model is to test for mediation by using each prior criterion variable as the first predictor in each successive hierarchical regression to control for any indirect effects transmitted through it, thus estimating only the residual direct effects of each subsequent predictor variable (Figueredo and Gorsuch 2007). As fewer historical data were available for the developmental instability (DI) variable, the sequential canonical cascade model was estimated exclusively on data spanning the years from AD 1825–1985. The protective omnibus Pillai-Bartlett trace test for the entire sequential canonical analysis model was statistically significant: V = 0.984, E = 0.57, 90%CI = (0.56, 0.58), F(3,104) = 2068.47, p <.0001.
Table 4 displays Cascade Equation 1, with time (YR) having a statistically significantly and negative direct effect on intergroup competition (IGC; supporting H1). The semipartial correlation coefficient is indicated by the symbol sR; Figure 2 shows this relation graphically, using the standardized (z) scores of both predictor and criterion variables. These results indicate that intergroup competition has been decreasing across time since AD 1825.
Table 4. Cascade Equation 1: Hierarchical Regression for IGC with YR from AD1825–1994
| Predictor | sR | C.I. (90%) | F | df1, df2 | p |
| YR | -0.19* | -0.36,0.01 | 3.77 | 1, 106 | 0.05 |
*p <.05
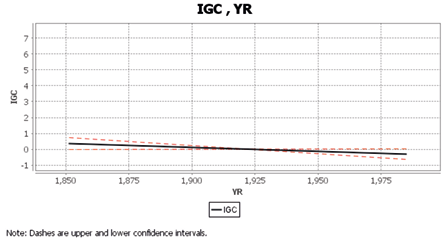
Fig. 2. Hierarchical Regression of the direct effect (semipartial correlation) of YR on IGC (AD 1825–1985).
Table 5 displays Cascade Equation 2, in which IGC had a statistically significant and negative direct effect on developmental instability (DI; supporting H2), while YR had a statistically significant and positive residual direct effect on DI (supporting H3). The semipartial correlation coefficient is indicated by the symbol sR; Figures 3 and 4 illustrate these relations graphically, using the standardized (z) scores of both predictor and criterion variables. These all indicate that DI is reduced by higher levels of intergroup competition, which Cascade Equation 1 shows to be declining, but that DI has otherwise been increasing through time since AD 1825.
Table 5. Cascade Equation 2: Hierarchical Regression for DI with IGC and YR from AD1825–1985
| Predictor | sR | C.I. (90%) | F | df1, df2 | p |
| IGC | -0.33* | -0.49, -0.14 | 22.89 | 1, 105 | <0.0001 |
| YR | 0.63* | 0.50, 0.74 | 86.23 | 1, 105 | <0.0001 |
*p <.05

Fig. 3. Hierarchical Regression of the residual direct effect (semipartial correlation) of IGC on DI (AD 1825–1985).
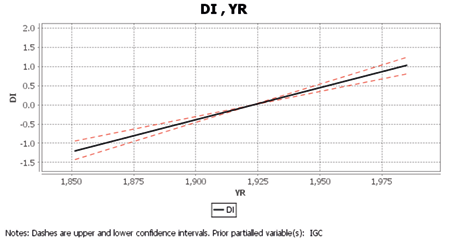
Fig. 4. Hierarchical Regression of the residual direct effect (semipartial correlation) of YR on DI, statistically controlled for the effect of IGC (AD 1825–1985).
Table 6 displays Cascade Equation 3, wherein DI had a statistically significantly and negative direct effect on Asabiyyah (supporting H4), while IGC had a statistically significantly and positive residual direct effect on Asabiyyah (supporting H5), and YR had a statistically significant and negative residual direct effect on Asabiyyah (supporting H6). The semipartial correlation coefficient is indicated by the symbol sR; Figures 5, 6, and 7 show these relations graphically, using the standardized (z) scores of both predictor and criterion variables. These all indicate that Asabiyyah is reduced by higher levels of developmental instability, which Cascade Equation 2 has shown to be rising, but that Asabiyyah increases with higher levels of intergroup competition, which Cascade Equation 1 shows to be declining, and Asabiyyah has otherwise been decreasing over time since AD 1825.
Table 6. Cascade Equation 3: Hierarchical Regression for ASABIYYAH with DI, IGC, and YR from AD1825–1985
| Predictor | sR | C.I. (90%) | F | df1, df2 | p |
| DI | -0.61* | -0.72,-0.48 | 543.44 | 1, 104 | <0.0001 |
| IGC | 0.06* | -0.25,0.14 | 4.75 | 1, 104 | 0.03 |
| YR | -0.74* | -0.82,-0.64 | 785.70 | 1, 104 | <0.0001 |
*p <.05

Fig. 5. Hierarchical Regression of the direct effect (semipartial correlation) of DI on ASABIYYAH (AD 1825–1985).

Fig. 6. Hierarchical Regression of the residual direct effect (semipartial correlation) of IGC on ASABIYYAH, statistically controlled for the effect of DI (AD 1825–1985).

Fig. 7. Hierarchical Regression of the residual direct effect (semipartial correlation) of YR on ASABIYYAH, statistically controlled for the effect of DI and IGC (AD 1825–1985).
Consistent with predictions from both the SEAM and the evoked culture model, the temporal decline in the Asabiyyah factor is independently predicted by both the (declining) intergroup competition and (rising) developmental instability factors. Critically, the developmental instability measure seems to substantially mediate the impact of declining intergroup competition on the Asabiyyah factor, consistent with the hypothesized impact of diminished intergroup competition on mutation accumulation, as predicted by SEAM. Furthermore, these results demonstrate a novel application of Haidt’s moral foundations theory to the elucidation of culturomic shifts and their determinants. We also note that the decline in Asabiyyah noted here is consistent with the work of Younes and Ulf-Dietrich (2019), who, in employing Google Ngram Viewer, found indications of a general decrease in collectivistic religious expression across multiple languages between 1900 and 2000, with (temporary) reversals to this trend having occurred during times of conflict (e.g., World War II).
The small and positive residual direct effect of intergroup competition on the Asabiyyah factor, after statistically controlling for that of developmental instability, was minimally consistent with the predictions of the evoked culture model. This model would hypothesise that declining intergroup competition might serve as a direct driver of changes in preferences from group-oriented to individual-oriented textual expressions of moral psychology. As discussed in the introduction, the process by which this path-dependency arises might relate to diminished levels of intergroup competition evoking pre-existing evolved psychological mechanisms that adaptively upregulate preferences for individualizing morality, with more fairness- and harm-avoidance-oriented moral expressions being more adaptive under conditions of intergroup peace. This process might also establish a selective context in which, via gene-culture coevolution, rapid selection can take place favoring the fitness of genotypes that predispose toward the development of these individualizing moral foundations. Such selection can conceivably even act over relatively short periods of time[5]
Further, the measurement model for the latent structure of Asabiyyah was not meaningfully confounded by temporal autocorrelations, which were found to be of negligible magnitudes, nor were the lexicographically convergent results confounded with the age of the words sampled, which is significant as age has been found in previous work on Ngram viewer to be a significant predictor of temporal changes in the utilization frequencies of words (Woodley of Menie et al. 2015). The developmental instability factor may therefore capture changes in the strength of negative selection on indicators that may serve as proxy measures of disturbed patterns of social epistasis. Moreover, evidence for this mediational pathway strengthens the case for lexicographic moral foundation measures serving as diachronic indicators of either positive or negative social epistasis.
These findings also have relevance for ongoing debates in evolutionary psychology concerning the possibility that certain levels of trait expression might be maladaptive – meaning that they stem from some process that is leading to long-term reductions of fitness. A related debate concerns the possibility that the extremely low fertility rates characteristic of post-demographic-transition Western populations, in particular, might be maladaptive in so far as their fertility is at sub-replacement levels. Arguments have been made to the effect that this consequence of the demographic transition merely reflects changes in patterns of bioenergetic investments stemming from adaptive, developmentally mediated transitions into slower life history, and specifically a regime of significantly diminished child, infant, and general mortality. Based on this alternative argument, lower individual-level fertility might therefore be “paid for” in other ways by changes in such patterns of investments resulting in for example, greater somatic persistence (longevity) and greater allocations of effort into certain communitarian domains (e.g., Colleran 2016).
These results should be interpreted with caution, insofar as some may be inclined to read them as suggesting that group-oriented ideologies are simply “better,” in terms of their effects on group fitness, than their individual-oriented counterparts. It is becoming increasingly apparent, from real-world evidence, that this is flatly incorrect, however. In the contemporary Western context at least, the adoption of extreme ideologies, including and perhaps especially group-oriented ones, for instance Communism on the left and neo-Nazism on the right, seems to be uniquely attractive to those whom criminologists would term “socially deviant” individuals. Such individuals are at elevated risk of a number of undesirable outcomes, such as engagement in crime (including violent crime and terrorism) and low social status. The organized actions of these individuals often seriously harm, particularly through violence, the welfare of the broader populations in which they are embedded (Institute for Economics and Peace 2020) and occasion the rapid dissolution of the movements with which they are associated. Even if it is true, as these results suggest, that genetic change in Western populations has been favoring the rise of excessively individualistic (from a group-fitness perspective) values, it does not follow from this that all groupish alternatives are functional or in some way desirable. Indeed, it could be that genetic and social-epistatic dynamics have taken long-standing individualistic tendencies in Western populations to problematic levels, while also giving rise to damaging group-oriented efforts to “correct” these developments. Pathological manifestations of extreme and groupish political and social ideologies obviously are nothing new. The especially catastrophic results, in moral and biological terms, of Communism and Nazism in the twentieth century speak for themselves – and it is hardly surprising that the intellectual heirs of these movements clearly share psychological traits widely considered to be highly socially undesirable (Costello et al. 2022; Moss and O’Connor 2020 a,b). Although the groupish social and political arrangements of premodern societies in the more distant past across the world were hardly liberal, and involved various moral evils, the disastrous totalitarianisms of recent history, and their contemporary ideological progeny, suggest the appearance of uniquely perverse extremisms. It must be stressed that all authors on the current paper unequivocally oppose extremist political views.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and material
Google Ngram Viewer data can be obtained from the following url: https://books.google.com/ngrams. The lexical items used as the basis for constructing diachronic measures of moral foundations were obtained from Graham, Haidt, and Nosek’s (2009) moral foundations dictionary. The diachronic data used here have been made publicly available in Hertler et al. (2020), Woodley of Menie et al. (2017b), and from and sources contained therein.
Code availability
All code will be made available upon request.
Ethics approval
Not applicable.
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Consent to participate
Not applicable.
Authors’ contributions
MWOM, MAS, and MPA drafted the manuscript. AJF, MPA, and MJ prepared the data analyses. All authors approved the final version.
Bachmann SO, Cross E, Kalbassi S, Sarraf MA, Woodley of Menie MA, Baudouin SJ. 2018. Protein pheromone MUP20/Darcin is a vector and target of indirect genetic effects in mice. bioRxiv 1–20.
Bolt J, Inklaar R, de Jong H, van Zanden JL. 2018. Rebasing ‘Maddison’: New income comparisons and the shape of long-run economic development. GGDC Research Memorandum 174.
Budnik A, Henneberg M. 2017. Worldwide increase of obesity is related to the reduced opportunity for natural selection. PLOS ONE 12:e0170098.
Colleran H. 2016. The cultural evolution of fertility decline. Phil Trans R Soc B 371:20150152.
Costello TH, Bowes SM, Stevens ST, Waldman ID, Tasimi A, Lilienfeld SO. 2022. Clarifying the structure and nature of left-wing authoritarianism. J Pers Soc Psychol 122:135–170.
Cross ESR. 2019. Investigation of social olfaction in a Neuroligin 3 Knockout mouse model. Unpublished doctoral dissertation, School of Biosciences, Cardiff University, UK.
Curzan A. 2009. Historical corpus linguistics and evidence of language change. In: A Luedeling and M Kytö, editors. Corpus linguistics. Berlin, Gruyter. 1091–1108.
Darwin C. 1871. The descent of man, and selection in relation to sex. London: John Murray.
Dawkins R. 1981. Selfish genes in race or politics. Nature 289:528.
Del Giudice M. 2012. The twentieth century reversal of pink-blue gender coding: a scientific urban legend? Arch Sex Behav 41:1321–1323.
Figueredo AJ, Gorsuch R. 2007. Assortative mating in the Jewel wasp: 2. Sequential canonical analysis as an exploratory form of path analysis. J Ariz-Nev acad sci 39:59–64.
Figueredo AJ, McKnight PE, McKnight KM, Sidani S. 2000. Multivariate modeling of missing data within and across assessment waves. Addiction 95 Suppl 3:361–380.
Figueredo AJ, Peñaherrera-Aguirre M, Fernandes HBF, Lomayesva SL, Woodley of Menie MA, Hertler SC, Sarraf MA. 2019a. The Ecology of Empire: The dynamics of strategic differentiation-integration in two competing Western European biocultural groups. PLS 38:210–225.
Figueredo AJ, Peñaherrera-Aguirre M, Fernandes HBF, Lomayesva SL, Woodley of Menie MA, Herter SC, Sarraf MA. 2019b. War and Peace: A diachronic social biogeography of life history strategy and between-group relations in two Western European populations. J Methods Meas Soc Sci 10:36–75.
Gorsuch RL. 1983. Factor Analysis (2nd ed.). Hillsdale, NJ: Lawrence Erlbaum Associates.
Gorsuch RL. 2016. UniMult: For univariate and multivariate data analysis. Altadena, CA: UniMult, Inc. Available at: https://unimult.000webhostapp.com/ [Accessed 30 May 2022].
Graham J, Haidt J, Nosek BA. 2009. Liberals and conservatives rely on different sets of
moral foundations. J Pers Soc Psychol 96:1029–1046.
Greenfield PM. 2013. The changing psychology of culture from 1800 through 2000. Psychol Sci 24:1722–1731.
Graham J, Nosek BA, Haidt J, Iyer R, Koleva S, Ditto PH. 2011. Mapping the Moral Domain. J Pers Soc Psychol 101:366–385.
Haidt J. 2007. Moral psychology and the misunderstanding of religion. Edge. Available at: https://www.edge.org/conversation/jonathan_haidt-moral-psychology-and-themisunderstanding-of-religion [Accessed 30 May 2022].
Haidt J. 2008. Morality. Perspect Psychol Sci 3:65–72.
Haidt J. 2012. The righteous mind: Why good people are divided by politics and religion. New York: Vintage Books.
Haidt J. 2016. Are moral foundations heritable? Probably. The Righteous Mind. Available at: https://righteousmind.com/are-moral-foundations-heritable-probably/#:~:text=Are%20moral%20foundation%20scores%20heritable,heritable%20nor%20stable%20over%20time [Accessed 30 May 2022].
Hamilton WD. 1971. Selection of selfish and altruistic behavior in some extreme models. In: JF Eisenberg, WS Dillon, editors. Man and beast: Comparative social behavior. Smithsonian Institution Press, Washington DC, USA, 57–91.
Hertler S, Figueredo AJ, Peñaherrera Aguirre M. 2020. Multilevel Selection: Theoretical Foundations, Historical Examples, and Empirical Evidence. New York, NY: Palgrave Macmillan.
Hills TT, Adelman JS. 2015. Recent evolution of learnability in American English from 1800 to 2000. Cognition 143:87–92.
Fieder M, Huber S. 2018. Political attitude and fertility: Is there a selection for the political extreme? Front Psychol 9:2343.
Ibn Khaldun AA-R. 1377/1969. The Muqaddimah: An Introduction to History. NJ Dawood, editor. Princeton, NJ: Princeton University Press.
Institute for Economics and Peace. 2020. Global terrorism index 2020: Measuring the impact of terrorism. National Consortium for the Study of Terrorism and Responses to Terrorism. Available at: https://visionofhumanity.org/wp-content/uploads/2020/11/GTI-2020-web-1.pdf
Kalbassi S, Bachmann SO, Cross E, Roberton VH, Baudouin SJ. 2017. Male and female mice lacking Neuroligin-3 modify the behavior of their wild-type littermates. eNeuro 4:e.0145–17.2017.
Kimmerle EH, Jantz RL. 2006. Secular trends in craniofacial asymmetry studied by geometric morphometry and generalized Procrustes methods. In: DE Slice, editor. Modern morphometrics in physical anthropology. Springer, 247–263.
Komlos J, Brabec M. 2010. The trend of mean BMI values of US adults, birth cohorts 1882—1986 indicates that the obesity epidemic began earlier than hitherto thought. Am J Hum 22:631–638.
Kondrashov AS. 2017. Crumbling genome: The impact of deleterious mutations on humans. Hoboken: Wiley Blackwell.
Ladle RJ, Correia RA, Do Y, Joo G-J, Malhando ACM, Proulx R, Jepson P. 2016. Conservation culturomics. Front Ecol Environ 14:269–275.
Linksvayer TA. 2007. Ant species differences determined by epistasis between brood and worker genomes. PLOS ONE 2:e994.
Lynch M. 2016. Mutation and human exceptionalism: our future genetic load. Genetics 202:869–875.
Markow TA. 1992. Human handedness and the concept of developmental stability. Genetica 87:87–94.
McKnight PE, McKnight KM, Sidani S, Figueredo AJ. 2007. Missing Data: A Gentle Introduction. New York, NY: Guilford Press.
McManus IC, Moore J, Freegard M, Rawles R. 2010. Science in the making: right hand, left hand. III: Estimating historical rates of left-handedness. Laterality 15:186–208.
Michel JB, Shen YK, Aiden AP, Veres A, Gray MK, Aiden EL. 2011. Quantitative analysis of culture using millions of digitized books. Science 331:176–182.
Moss JT, O’Connor PJ. 2020a. Political correctness and the alt-right: The development of extreme political attitudes. PLOS ONE 15:e0239259.
Moss JT, O’Connor PJ. 2020b. The Dark Triad traits predict authoritarian political correctness and alt-right attitudes. Heliyon 6:e04453.
Ritz C, Spiess AN. 2008. qpcR: an R package for sigmoidal model selection in quantitative real-time polymerase chain reaction analysis. Bioinformatics 24:1549–1551.
Rohatgi A. 2017. WebPlotDigitizer. Available at: https://apps.automeris.io/wpd/ [Accessed 30 May 2022].
Roivainen E. 2014. Changes in word usage frequency may hamper comparisons of vocabulary skills: an Ngram analysis of Wordsum, WAIS and WISC test items. J Psychoeduc Assess 32:83–87.
Roser M, Ritchie H, Ortiz-Ospina E. 2013. World population growth. Available at: https://ourworldindata.org/world-populationgrowth [Accessed 30 May 2022].
Rühli F, Henneberg M. 2017. Biological future of humankind – Ongoing evolution and the impact of recognition of human biological variation. In: M Tibayrenc and FJ Ayala, edtors. On human nature: Biology, psychology, ethics, politics, and religion. London, UK: Elsevier. 263–275.
SAS Institute Inc. 2015. Base SAS 9.4 Procedures guide: Statistical Procedures (5th ed.). Cary, NC: SAS Institute Inc. Available at: http://support.sas.com [Accessed 30 May 2022].
Smith KB, Alford JR, Hibbing JR, Martin NG, Hatemi PK. 2016. Intuitive ethics and political orientations: testing moral foundations as a theory of political ideology. Am J Pol Sci 61:424–437.
Tooby J, Cosmides L. 1992. The psychological foundations of culture. In: J.H. Barkow, L. Cosmides, J. Tooby, editors. The adapted mind: Evolutionary psychology and the generation of culture.Oxford University Press, 19–136.
Van Valen L. 1962. A study of fluctuating asymmetry. Evolution 16:125–142.
Woodley of Menie MA, Fernandes HBF. 2016. The secular decline in general intelligence from decreasing developmental stability: theoretical and empirical considerations. Pers Individ Differ 92:194–199.
Woodley of Menie MA, Fernandes HBF, Figueredo AJ, Meisenberg G. 2015. By their words ye shall know them: Evidence of genic selection against general intelligence and concurrent environmental enrichment in vocabulary usage since the mid 19th century. Front Psychol 6:361.
Woodley of Menie MA, Figueredo AJ, Sarraf MA. 2019. Slowing life history (K) can account for increasing microinnovation rates and GDP growth, but not macroinnovation rates, which declined following the end of the Industrial Revolution. BBS 42, e213.
Woodley of Menie MA, Figueredo AJ, Sarraf MA, Hertler S, Fernandes HBF, Peñaherrera Aguirre M. 2017b. The Rhythm of the West: A biohistory of the modern era, AD 1600 to present. J Soc Political Econ Stud. Monograph Series 37. Washington, DC: Council for Social and Economic Studies.
Woodley of Menie MA, Kanazawa S, Pallesen J, Saraf MA. 2020. Paternal age is negatively associated with religious behavior in a post-60s but not a pre-60s US birth cohort: Evidence for the Social Epistasis Amplification Model. J Relig Health 59:2733–2752.
Woodley of Menie MA, Saraff M, Pestow R, Fernandes HBF. 2017a. Social epistasis amplifies the fitness costs of deleterious mutations, engendering rapid fitness decline among modernized populations. Evol Psychol Sci 3:181–191.
Younes N, Ulf-Dietrich R. 2019. Guideline for improving the reliability of Google Ngram studies: Evidence from religious terms. PLOS ONE 14:e0213554.
| Abandon | Brutality | Cruelty | Disgust | Exploiting | Hierarchy | Insider | Loyalty | Position | Respect | Solidarity | Unchaste |
| Abstemiousness | Cadre | Crusher | Dishonest | Exploits | Holiness | Insubordination | Maiden | Preference | Respected | Spurn | Uncleanliness |
| Abstention | Care | Damage | Disloyalty | Fair | Holy | Insurgent | Member | Prejudice | Respectfulness | Spy | Unequal |
| Abstinence | Caring | Debasement | Disobedience | Fair play | Homeland | Integrity | Miscreant | Preserve | Respects | Stain | Unfairness |
| Abuse | Caste | Debauchery | Disproportion | Fairly | Homologous | Intemperate | Modesty | Pristine | Rights | Status | Unfaithful |
| Adultery | Celibacy | Deceiver | Disrespect | Fairmindedness | Honesty | Jilter | Mother | Profanity | Riot | Sterility | Unison |
| Agitation | Chastity | Decency | Dissent | Fairness | Honorable | Joint | Mothering | Profligate | Ruin | Stomp | Uniter |
| Alienate | Class | Defector | Dissident | Familial | Humble | Justice | Motherland | Promiscuity | Sacredness | Subversion | Unjustness |
| Ally | Cleanliness | Defense | Dissociate | Families | Hurt | Justification | Mothers | Prostitute | Safety | Suffering | Unprejudicedness |
| Amity | Clique | Defiance | Egalitarianism | Family | Illegality | Justness | Mutinous | Protection | Saint | Supremacy | Unscrupulous |
| Annihilation | Cohort | Defiler | Empathy | Father | Immaculate | Kill | Nation | Protest | Security | Taint | Upright |
| Apostasy | Collective | Denounce | Endangerment | Favoritism | Immigrant | Killed | Nonconformist | Pureness | Sedition | Tarnish | Violence |
| Apostate | Command | Depravity | Enemy | Fellowship | Impair | Killer | Obeyance | Purity | Segregation | Terrorism | Virgin |
| Attack | Communal | Desecration | Equable | Fight | Impartiality | Killing | Obscenity | Rank | Sequester | Together | Virginal |
| Austerity | Commune | Deserted | Equality | Filth | Impiety | Kills | Obstruct | Ravage | Shelter | Tolerant | Virginity |
| Authority | Communism | Deserter | Equity | Foreigner | Impious | Law | Oppose | Reasonable | Shield | Tradition | Virgins |
| Balance | Community | Deserting | Equivalent | Gross | Imposter | Lawfulness | Order | Rebel | Sickness | Traitor | Virtuous |
| Benefit | Compassion | Destroy | Evenness | Group | Indecency | Lawlessness | Peace | Reciprocity | Sin | Tramp | Wanton |
| Betrayal | Compliance | Detriment | Excluder | Guard | Individual | Lax | Permission | Refined | Sinfulness | Trashy | War |
| Bias | Comrade | Devotee | Exclusion | Guild | Inequitable | Leader | Permit | Refuse | Sinned | Treachery | Warlord |
| Bigotry | Constant | Dirt | Exploit | Harm | Ingroup | Legality | Pervert | Remonstrate | Sinner | Treason | Warring |
| Blemish | Contagion | Discrimination | Exploitation | Harm | Injustice | Lewdness | Piety | Renegade | Sinning | Unadulterated | Wars |
| Bourgeoisie | Control | Disease | Exploited | Heretic | Innocent | Limpid | Pious | Repulsion | Sins | Unbiasedness | Wholesomeness |
S1. Abridged list of Moral Foundation words collected from Haidt (2012) and the online version of the Moral Foundations Questionnaire.


Received: 2022.02.15; Revised: 2022.04.19; Accepted: 2022.04.25