Introduction
Although the current understanding of human evolution away from the common ancestor with apes prefers the “branching tree model” there was only one way to produce presently living humans: by subjecting some late Miocene hominid to evolutionary forces that ultimately produced the reader of this text and contemporaneous human conspecifics. While all published trees by convention are shown branching upward from our Miocene ancestor to ourselves, they are imaginary (a fact that can be inferred from their very variety) and very few of them are validated by anything but repetitive group-think. An example of such a tree is in Figure 1.
Fig. 1. An example of an “evolutionary tree” of humans. Authors own drawing not based on any specific considerations of links between “fossil species”. Just an example.
Instead, it is preferable to reverse perspective and look backward through the telescope that connects the broad observable reality of our current species with – from any reasonable evolutionary perspective – its logically essential and paleontologically documented (Galik et al. 2004) antecedent populations.
The history of this subject, as history usually is with its failures and achievements, is what we are trying to explain here. Although any particular explanation may be only hypothetical and uncertain, it is absolutely necessary that the evolutionary process producing modern humans actually happened – look in the mirror if uncertain. With the current branching model of hominin evolution misrepresenting members of various populations such as Neandertals and even Flores “hobbits”, as purported failures to become “moderns” (in the sense of morphology and behaviour rather than their time of existence), it is unavoidably essential that there was a direct ancestor-descendant line that, proceeding through the copulations and births in every 250,000 antecedent generations, produced modern humans from some Miocene/Pliocene ancestral population. Differential reproduction in each of those 250,000 generations enabled natural selection to enhance biological processes that underlie complex cognition of modern human individuals. Interaction of members of each generation with their surroundings and with other members of their populations influenced epigenetic processes shaping the course of individual ontogenies, especially their early parts: infancy, childhood and adolescence, and resulted in altered natural environments and social structures with which all our ancestors had to interact.
The fossil record of hominin morphologies and the corresponding archaeological records of results of hominin behaviours are all imperfect due to the vagaries of taphonomy and geological preservation. Nonetheless, where fossil or archaeological sampling has provided reasonable sample sizes of skeletal remains or artefacts, it can be seen that the evidence shows improvements in bodily characteristics or technological abilities of some hominins increasing at a double exponential rate through the last few million years. The characteristic investigated should be measured on an interval (continuous) scale.
The general form of the double exponential equation is
where y – a dependent variable, x – an independent variable (e.g. time), a, b, constants.
As an example, for the increase in hominin cranial capacity representing the increase in body size the equation is (Fig. 2)
Where CC is measured in millilitres (ml) and DATE in thousands of years (ka).
The double exponential rate of increase is the basic formal characteristic of self-amplifying systems.
Double exponential regression of hominin cranial capacities found until the end of Pleistocene (N=233) on their dates has a strong fit to actual data (r=0.92). Variance of actual capacities around the fitted line (stdev=164) does not differ significantly from the variance of brain size among the one species of modern humans (stdev=157, F=1.09; Henneberg 1990).
Cranial capacity is easy to quantify, while technological advances or other traces of behaviours do not lend themselves easily to reliable quantification (Levendis et al. 2019). This evidential constraint, however, does not mean that cultural innovations are not a result of self-amplifying feedback relations. The following basic behavioural characteristics often are mentioned, as being involved in the operation of the human system of positive feedbacks (Bielicki 1969; Tobias 1981; Henneberg 1992; McKee 2017): erect bipedalism, food procurement, technologies, symbolic communication, extended childhood, sexual behaviours, social organisation, cognitive abilities. Erect bipedalism, however acquired, became a very energetically efficient way of locomotion on the ground and in shallow waters, freeing up the upper extremities. These could be then used for transporting food, weapons/tools, raw materials and children in a position facing the carrier. Similar to trunks of proboscideans, this shift extended the range of individuals’ manipulation of the environment, and added to it increased interaction between care-givers and offspring. The erect posture liberated hands that then could manipulate various objects including foodstuffs, tools/weapons, fuels and building materials even during locomotion and without use of teeth and mouths; mouth then could be used to send voice signals improving interindividual communication. Such effects of erect bipedalism clearly enhanced food acquisition and food sharing, and facilitated development of technologies, rearing of offspring, symbolic communication that in their own way interacted with the enhanced mental abilities and social organisation. Concomitant changes in sexual behaviours improved cooperative parenting and cooperation in food acquisition (to a large extent multi-person foraging, scavenging and hunting). Changed sexual behaviours benefitted from ovulatory crypsis (Lovejoy 2009) allowing more frequent, nearly continuous, female sexual receptivity, promoting monogamous unions with males that might also contribute to cooperative parenting.
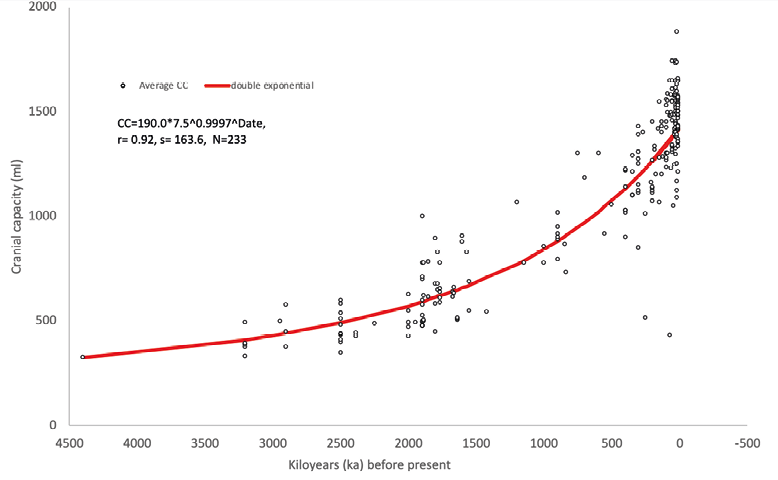
Fig. 2. Illustration of the quantitative operation of the self-amplifying feedbacks on human body size as represented by the volume of the cranial cavity. Data taken from De Miguel and Henneberg (2001) and Henneberg and De Miguel (2004) supplemented by data on new finds (Jacob et al. 2006; Berger et al. 2010; De Ruiter et al. 2019).
Collaborative parenting provided an opportunity to prolong childhood, a period of rapid and extensive learning. A prolonged childhood period enabled individuals to acquire knowledge and skills needed to use more sophisticated technologies and to participate in more intricate social structures. All these changes eventually required more learning thus demanding a further extension of the period of childhood. Improved technologies produced more efficient ways of food procurement resulting in greater access to nutrients. This interactive complex resulted in larger number of individuals being able to live together and cooperate in child rearing and food procurement but complicated social relations that had to be modified. Improved social relations enhanced collaborative parenting and coordinated food procurement that resulted in increased group sizes requiring further development of social organisation. Such improvement was made possible by developed mental capacities and inter-individual communication (language). Improved mental abilities and more efficient (eg. symbolic) communication contributed to better social organisation that enhanced production and use of new technologies and more abundant food acquisition. Greater food abundance allowed larger group size, and so on and so on, iteratively through vast stretches of time… The scheme has many uncertain details but its general heuristic value is undeniable (Figure 3).
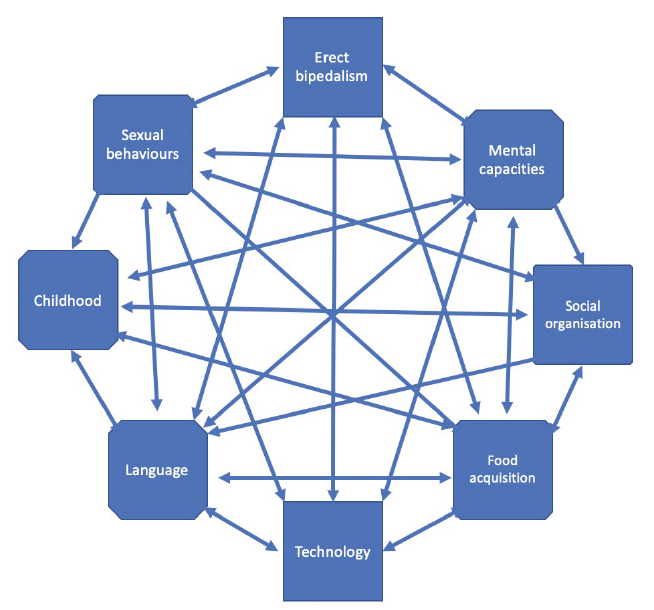
Fig. 3. A schematic representation of autocatalytic feedbacks operating during human evolution.
Evidence for the operation of this set of self-amplifying feedbacks extends far beyond the simple example of cranial capacity increase. Initial low population density of hominins, evidenced by just a few fossil finds from the Miocene and Pliocene, expanded exponentially into Pleistocene leaving fossil evidence everywhere in the Old World, even if one discounts obvious taphonomic limitations and natural vagaries of preservation. The initial scarcity of (mostly stone) artefacts of Lomekwi and of Oldovan technologies in time flourished into Acheulian industries of Lower and Middle Pleistocene and abundance of human made objects, not only technologically useful but also decorative, and into natural surfaces’ alterations (e.g. “cave art”) of the Upper Palaeolitic and climaxed in the management of plants, animals, soils, watercourses and building materials of the Neolithic. The abundance of human ways to manage the environment that occurred in the Neolithic, altered direction of some feedbacks so that, for example, increasing body and brain size as an effect of better food acquisition, producing collateral effects of greater protection against attacks of predators and imoproved hunting efficiency, later reversed towards reduction in body size (gracilisation, Debec 1960; Schwidetzky 1962) and brain size (Henneberg 1988; Henneberg and Steyn 1993) to lower metabolic costs of body mass in the situation of strong group protection and more reliable, though quantitatively limited, food supply from agricultural activities.
The mechanism described here produced, at any time it operated, a variety of morphologies and behaviours that still were capable of engaging in the set of positive feedbacks, and continued towards characteristics that we now perceive as those of modern humans. The rates of morphological and behavioural changes may have varied at different periods, but continuity of change has not been interrupted in the set of our ancestral populations. Therefore, search for a single moment in time when ‘modernity’ appeared, or for the individual who could be the earliest holotype of modern Homo sapiens is a futile exercise because human variation in time and geographical space is shaped by ongoing processes, not a single act of … creation… by forces, however natural.
Authors’ contributions
Both authors contributed equally to this paper’s contents.
Conflict of interest
Authors declare no conflict of interests.
References
Bielicki T. 1969. Niektóre związki zwrotne w procesie ewolucji Hominidae. [In Polish]. Eng: Deviation-amplifying cybernetic systems and hominid evolution]. Mat. i Prace Antrop 77:3–60.
Berger LR, De Ruiter DJ, Churchill SE, Schmid P, Carlson KJ, Dirks PHGM, Kibii JM. 2010. Australopithecus sediba: a new species of Homo-like australopith from South Africa. Science 328(5975):195–204. https://doi.org/10.1126/science.1184944
Debec GF. 1960. Certain aspects des transformations somatiques de l’Homo sapiens. Communiques de la delegation soviétique au Ve Congrès International des Sciences Anthropologiques, Moscow. p 25.
De Miguel C, Henneberg M. 2001. Variation in hominid brain size: How much is due to method. Homo 52(1):3–58. https://doi.org/10.1078/0018-442X-00019
De Ruiter DJ, Laird MF, Elliott M, Schmid P, Brophy J, Hawks J, and Lee R. Berger LR. 2019. Homo naledi cranial remains from the Lesedi chamber of the rising star cave system, South Africa. J Hum Evol 132:1–14. https://doi.org/10.1016/j.jhevol.2019.03.019
Galik K, Senut B, Pickford M, Gommery D, Treil J, Kuperavage AJ, and Robert B. Eckhardt RB. 2004. External and internal morphology of the BAR 1002’00 Orrorin tugenensis femur. Science 305(5689):1450–53. https://doi.org/10.1126/science.1098807
Henneberg M. 1988. Decrease of human skull size in the Holocene. Hum Biol 60(3):395–405.
Henneberg M. 1990. Brain size/body weight variability in Homo sapiens: consequences for interpreting hominid evolution. Homo 39:121–30.
Henneberg M. 1992. Continuing human evolution: bodies, brains and the role of variability. Trans Roy Soc S Africa 48(2):159–82. https://doi.org/10.1080/00359199209520260
Henneberg M, de Miguel C. 2004. Hominins are a single lineage: brain and body size variability does not reflect postulated taxonomic diversity of hominins. Homo 55:21–37. https://doi.org/10.1016/j.jchb.2004.03.001
Henneberg M, Steyn M. 1993. Trends in cranial capacity and cranial index in Subsaharan Africa during the Holocene. Am J Human Biol 5:473–79. https://doi.org/10.1002/ajhb.1310050411
Jacob T, Indriati E, Soejono RP, Hsü K, Frayer D, Eckhardt RB, Kuperavage AJ, Thorne A, Henneberg M. 2006. Pygmoid Australomelanesian Homo sapiens skeletal remains from Liang Bua, Flores: population affinities and pathological abnormalities. Proc Natl Acad Sci 103(36):13421–6. https://doi.org/10.1073/pnas.0605563103
Levendis J, Eckhardt RB, Block W. 2019. Evolutionary psychology, economic freedom, trade and benevolence. Review of Economic Perspectives 19(2):73–94. https://doi.org/10.2478/revecp-2019-0005
Lovejoy CO. 2009. Reexamining human origins in light of Ardipithecus ramidus. Science 326(5949):74–74e8. https://doi.org/10.1126/science.1175834
McKee JK. 2017. Correlates and catalysts of hominin evolution in Africa. Theory in Biosciences 136(3–4):123–40. https://doi.org/10.1007/s12064-017-0250-5
Schwidetzky I. 1962. Das grazilisierungsproblem. Homo 13:188–95.
Tobias PV. 1981. Evolution of Human Brain, Intellect and Spirit: University of Adelaide. The University of Adelaide, South Australia, Adelaide.


Received: 2021-05-12. Revised: 2022-01-28. Accepted: 2022-01-28